
产品名称: 稻壳酮A
英文名: Momilacton A
中文别名:
英文别名: Momilactone A
Cas 号: 51415-07-7
产品编码:BP5234
分子式: C20H26O3
分子量: 314.425
来源:
化合物类型: 萜类(Terpenoids)
纯度: 95%~99%
分析方法: HPLC-DAD or/and HPLC-ELSD
鉴定方法: 质谱(Mass), 核磁(NMR)
包装: 棕色小玻璃瓶,按客户需求包装。
存储: 贮存在避光密闭容器中,冷藏或者冷冻长期保存。
样品溶液最好临用新配。如果需要提前配制的话,最好分成独立包装冷冻保存(-20℃以下),临用前再取出解冻,通常可以保存2周。
可以满足克级以上大量需求,详情请咨询。
稻壳酮A和B是水稻抑制杂草的主要化感物质,其中稻壳酮B是至今发现最高效的天然除草剂之一, 具有很好的应用潜力。稻壳酮B可以从水稻根系释放到土壤中抑制周围稗草等植物的种子萌芽和生长。稻壳酮B的浓度大于3nmol·mL-1时就能抑制水芹和莴苣的根和胚轴生长, 同时稻壳酮A和B是水稻重要的植保素, 可有效抑制水稻病原菌比如稻瘟病菌(Magnaporthe grisea)等。此外, 稻瘟病菌感染可诱导水稻合成更多的稻壳酮。
越南科学家陈登春(音)教授和他在日本广岛大学的同事在稻壳中发现了抗癌物质。
这项题为《稻壳酮A和B抵抗急性早幼粒细胞白血病和多发性骨髓瘤细胞株的细胞毒性机制》的研究于今年10月发表在《癌症》杂志上。
报道称,这是第一个阐述稻壳酮A和稻壳酮B对急性早幼粒细胞白血病细胞HL-60和多发性骨髓瘤细胞株U266的细胞毒性机制的研究。
此外,这两种化合物还可能通过激活p-38蛋白以及破坏周期蛋白依赖性激酶(CDK1)和细胞周期蛋白B(Cyclin B1)形成的复合物,来诱导HL-60细胞周期的G2期阻滞。
这一发现将提供有关稻壳酮抗癌功能的新见解,从而可能成为未来基于稻壳酮的抗癌药物研究和开发的前提。
陈登春领导的广岛大学植物生理学和生物化学实验室是世界上少数可以从水稻等自然来源中提纯出稻壳酮的实验室之一。在2019年的一项研究中,该研究小组曾发现并成功地从30千克稻壳中提取了600毫克的稻壳酮A和稻壳酮B。
这位科学家说,日本国内外的许多制药公司正在与他的团队合作提取稻壳酮A和稻壳酮B,以用于研制免疫系统增强药物、抗癌药物、糖尿病药物及化妆品。
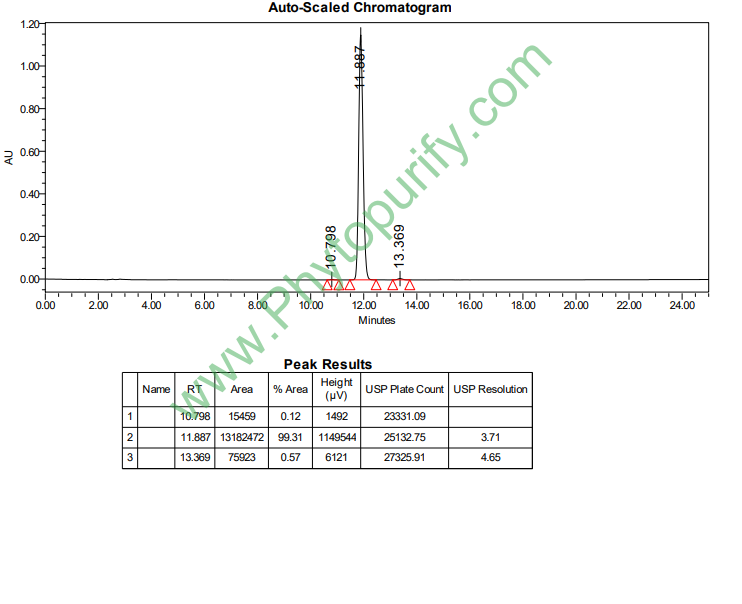
稻壳酮A的HPLC图谱
储存条件:短期保存 2~8℃,长期保存 -20 ~ -80℃
43.37
3.57
3.57
0.01
6.35
18.22
高
76.89
5.02
否
是
否
否
有
无
0.0
是
否
否
是

天然产物作为药物发现与农业化学的重要源泉,其独特的化学结构和生物活性长期以来吸引着研究者的目光。在众多具有生物活性的植物次生代谢产物中,来自水稻(Oryza sativa)及其近缘植物的稻壳酮类化合物(Momilactones)因其在植物防御和化感作用中的核心角色而备受关注。稻壳酮A(Momilacton A, CAS号:51415-07-7)是这一家族中最具代表性的成员之一,属于海螺烷二萜类化合物(pimarane-type diterpenoid),并具有独特的内酯结构。
稻壳酮A最初于1973年由日本科学家从水稻的稻壳中分离鉴定,并因其对稻瘟病菌(Magnaporthe oryzae)的强烈抑制作用而得名。随后的研究揭示,稻壳酮A不仅是水稻抵御病原微生物入侵的关键植保素(phytoalexin),还在水稻与其他植物(尤其是杂草)的化感相互作用中扮演着重要角色。这种双重功能——既是防御武器,又是生态竞争工具——使得稻壳酮A成为研究植物化学防御与生态适应性的理想模型分子。
近年来,随着对天然产物药理活性研究的深入,稻壳酮A的生物学功能已远远超越了植物保护领域。研究发现,稻壳酮A及其类似物对哺乳动物细胞表现出多种生物活性,包括抗炎、抗肿瘤、抗氧化以及神经保护等作用。这些发现为将这种古老的植物防御分子转化为治疗人类疾病的先导化合物提供了新的可能性。然而,尽管其植物生物学功能已被广泛研究,其作为潜在药物候选分子的成药性、药代动力学特性以及具体分子机制仍有待系统阐明。
本文旨在全面综述稻壳酮A的研究进展,涵盖其化学结构与理化性质、植物来源与提取方法、药理活性、作用机制与分子靶点、成药性评价与药代动力学特征,并探讨其在农业与医药领域的应用前景。通过整合多学科的研究成果,本文将为天然产物化学、植物生理学及药物开发领域的研究者提供一份系统、深入的参考资料。
稻壳酮A的化学结构属于海螺烷型二萜(pimarane-type diterpene),其核心骨架由四个异戊二烯单元(C20)构成,形成三环二萜体系。具体而言,其结构包含一个全氢菲(perhydrophenanthrene)骨架,并在C-8位和C-9位之间形成一个独特的γ-内酯环(γ-lactone),这是稻壳酮类化合物区别于其他二萜的关键结构特征。该内酯环由C-8位的羧基与C-9位的羟基脱水形成,赋予分子一定的刚性构象和化学反应活性。
稻壳酮A的分子式为C20H26O3,分子量为314.4250 g/mol。其结构中的官能团包括:一个α,β-不饱和酮(位于A环)、一个γ-内酯环(连接B环和C环)、以及多个手性中心。这些官能团的存在决定了其与生物靶标的相互作用模式。值得注意的是,稻壳酮A与稻壳酮B(Momilacton B)结构相似,区别仅在于B环上取代基的差异,这种细微的结构变化导致了生物活性的显著差异。
基于计算化学与实验测定,稻壳酮A的理化性质可归纳如下:
总体而言,稻壳酮A的化学结构决定了其兼具亲脂性和适度极性的特点,这使其在生物体内可能表现出复杂的分布和代谢行为。低水溶性和高BBB穿透性是成药性评价中需要重点关注的两个方面。
稻壳酮A最初从水稻(Oryza sativa)的稻壳中分离得到,因此得名。随后研究发现,稻壳酮A并非水稻所独有,而是广泛存在于禾本科(Poaceae)植物中,尤其是在稻属(Oryza)和稗属(Echinochloa)植物中含量较高。具体来源包括:
值得注意的是,稻壳酮A在植物体内的含量受多种因素调控,包括发育阶段、组织部位、环境胁迫(如干旱、盐碱、病原菌侵染)以及外源激素处理。例如,茉莉酸(jasmonic acid)和乙烯(ethylene)信号通路是调控稻壳酮A生物合成的关键上游信号。
鉴于稻壳酮A在植物组织中的含量通常较低(尤其是非胁迫条件下),其提取和纯化需要采用高效、特异性的方法。目前常用的提取流程包括以下几个步骤:
原料预处理:将新鲜或干燥的植物材料(如稻壳、水稻叶片)粉碎,过40-60目筛。为提高提取效率,可对原料进行脱脂处理(如使用正己烷或石油醚)以去除脂溶性杂质。
溶剂提取:稻壳酮A为中等极性化合物,通常采用极性有机溶剂进行提取。常用的溶剂体系包括:
提取方式可采用冷浸、超声辅助提取(UAE)或加速溶剂萃取(ASE)。超声辅助提取因其高效、温和的特点而被广泛采用。
液-液分配:将粗提物溶于水-有机溶剂(如正己烷、乙酸乙酯、正丁醇)中进行分级萃取。稻壳酮A主要富集于乙酸乙酯相中,可有效去除水溶性杂质和强脂溶性杂质。
柱色谱纯化:这是获得高纯度稻壳酮A的关键步骤。常用的色谱方法包括:
制备型高效液相色谱(Prep-HPLC):采用反相C18柱,以乙腈-水或甲醇-水为流动相,可得到纯度>98%的稻壳酮A。
结构鉴定:纯化后的化合物通过核磁共振波谱(NMR,包括1H-NMR、13C-NMR、HMBC、HSQC等)和高分辨质谱(HR-MS)进行结构确认。
近年来,随着绿色化学理念的推广,超临界流体萃取(SFE)和深共晶溶剂(DES)提取等新型方法也开始应用于稻壳酮A的提取,旨在提高效率、降低成本并减少有机溶剂的使用。
稻壳酮A最经典的药理活性体现在植物界。作为水稻的主要植保素,其在抵御稻瘟病菌(Magnaporthe oryzae)和纹枯病菌(Rhizoctonia solani)等病原真菌方面表现出显著活性。研究表明,稻壳酮A通过破坏真菌细胞膜的完整性、抑制孢子萌发和菌丝生长来发挥抗菌作用。其最小抑菌浓度(MIC)通常在10-50 μg/mL范围内,对某些专性寄生菌的活性尤为突出。
在化感作用方面,稻壳酮A是水稻抑制稗草等杂草生长的关键化学物质。水稻根系分泌的稻壳酮A可被邻近杂草吸收,通过干扰其细胞分裂、光合作用和激素信号通路来抑制生长。这一发现为开发基于天然产物的生物除草剂提供了重要思路。
近年来,稻壳酮A在哺乳动物细胞中的抗炎活性引起了广泛关注。体外实验表明,稻壳酮A能够显著抑制脂多糖(LPS)诱导的巨噬细胞(如RAW264.7细胞)中一氧化氮(NO)和前列腺素E2(PGE2)的产生。其机制涉及抑制诱导型一氧化氮合酶(iNOS)和环氧合酶-2(COX-2)的表达,以及下调促炎细胞因子如肿瘤坏死因子-α(TNF-α)、白细胞介素-6(IL-6)和白细胞介素-1β(IL-1β)的释放。
此外,稻壳酮A还可通过抑制核因子κB(NF-κB)和丝裂原活化蛋白激酶(MAPK)信号通路的激活来发挥抗炎作用。这些发现提示稻壳酮A可能对炎症性疾病(如关节炎、结肠炎)具有治疗潜力。
稻壳酮A对多种癌细胞系表现出细胞毒性作用。研究显示,其对肝癌细胞(HepG2)、乳腺癌细胞(MCF-7)、肺癌细胞(A549)和结肠癌细胞(HT-29)的增殖具有抑制作用,IC50值通常在10-50 μM范围内。机制研究表明,稻壳酮A可通过以下途径诱导癌细胞凋亡:
- 线粒体途径:上调Bax/Bcl-2比值,促进细胞色素c释放,激活caspase-9和caspase-3。
- 内质网应激:诱导GRP78、CHOP等蛋白表达,触发未折叠蛋白反应(UPR)。
- 细胞周期阻滞:将细胞周期阻滞于G2/M期,抑制有丝分裂进程。
值得注意的是,稻壳酮A对正常细胞(如人肝细胞L02)的毒性相对较低,表现出一定的选择性,这为其作为抗肿瘤先导化合物提供了安全性基础。
稻壳酮A还显示出一定的抗氧化活性。在DPPH和ABTS自由基清除实验中,其清除能力虽弱于维生素C,但优于许多其他二萜类化合物。此外,稻壳酮A可激活核因子E2相关因子2(Nrf2)信号通路,上调抗氧化酶如血红素加氧酶-1(HO-1)和醌氧化还原酶-1(NQO1)的表达,从而保护细胞免受氧化应激损伤。
在神经保护方面,稻壳酮A对谷氨酸诱导的HT-22海马神经元细胞损伤具有保护作用,可减少活性氧(ROS)积累和细胞凋亡。结合其高血脑屏障穿透性,稻壳酮A在阿尔茨海默病、帕金森病等神经退行性疾病的治疗中可能具有潜在价值。
此外,稻壳酮A还被报道具有抗病毒(如抑制流感病毒神经氨酸酶活性)、降血糖(通过激活AMPK信号通路)和免疫调节等活性。这些多样化的药理作用表明,稻壳酮A是一个具有多靶点作用特征的天然产物。
在植物系统中,稻壳酮A的作用机制与其作为化感物质和植保素的功能密切相关。研究表明,稻壳酮A可干扰靶标植物的激素信号网络,尤其是生长素(auxin)和油菜素内酯(brassinosteroid, BR)信号通路。
油菜素内酯信号通路:稻壳酮A被证实可抑制BR信号转导。BR信号始于细胞膜上的受体激酶BRI1(BRASSINOSTEROID INSENSITIVE 1)与其共受体BAK1(BRI1-ASSOCIATED RECEPTOR KINASE 1)的结合。稻壳酮A可能通过干扰BRI1-BAK1复合物的形成或抑制下游激酶BSK1(BR-SIGNALING KINASE 1)的磷酸化,从而阻断信号传递。这导致BSU1(BRI1 SUPPRESSOR 1)磷酸酶活性降低,进而使转录因子BZR1(BRASSINAZOLE RESISTANT 1)和BES1(BRI1-EMS-SUPPRESSOR 1)无法去磷酸化并入核,最终抑制BR响应基因的表达。这一机制解释了稻壳酮A为何能抑制植物细胞伸长和分裂。
生长素信号通路:稻壳酮A还影响生长素的极性运输和信号感知。它可能通过调节生长素外排载体PIN1(PIN-FORMED 1)的定位或活性,改变生长素在根尖和茎尖的分布梯度。此外,稻壳酮A可干扰生长素受体TIR1(TRANSPORT INHIBITOR RESPONSE 1)与AUX/IAA蛋白的相互作用,从而影响生长素响应因子ARF1(AUXIN RESPONSE FACTOR 1)的转录活性。这些作用共同导致靶标植物根生长抑制、叶片黄化等表型。
在哺乳动物细胞中,稻壳酮A的作用机制更为复杂,涉及多个信号通路和分子靶点:
初步的构效关系研究表明,稻壳酮A的γ-内酯环和α,β-不饱和酮结构是其生物活性所必需的。内酯环的开环或还原会显著降低其抗菌和抗炎活性。此外,C-8位的羧基和C-9位的羟基对于维持内酯环的稳定性和与靶蛋白的氢键相互作用至关重要。这些信息为后续的结构优化提供了重要指导。
基于前述理化性质,稻壳酮A的成药性可进行如下评价:
关于稻壳酮A在哺乳动物体内的药代动力学研究目前十分有限,主要基于计算预测和少量动物实验:
为克服稻壳酮A的低水溶性和潜在的低生物利用度,可考虑以下制剂策略:
- 纳米制剂:脂质体、聚合物纳米粒或固体脂质纳米粒可提高其溶解度和生物利用度。
- 环糊精包合物:β-环糊精或其衍生物可增加稻壳酮A的表观溶解度。
- 磷脂复合物:形成磷脂复合物可改善其脂溶性和跨膜能力。
- 前药设计:将内酯环开环形成羧酸酯前药,在体内经酶解再生为原药。
稻壳酮A最直接的应用前景在于农业领域。作为天然的化感物质和植保素,它有望被开发为:
- 生物农药:用于防治稻瘟病、纹枯病等水稻真菌病害,减少化学农药的使用。
- 生物除草剂:基于其抑制杂草生长的活性,开发环境友好型除草剂。特别是针对稗草等抗性杂草,稻壳酮A可能提供新的作用机制。
- 植物生长调节剂:通过调控BR和生长素信号通路,稻壳酮A或其类似物可用于调节作物株型、根系发育和抗逆性。
在医药领域,稻壳酮A的多种药理活性为其开发提供了多个方向:
- 抗炎药物:针对慢性炎症性疾病(如类风湿关节炎、炎症性肠病),稻壳酮A或其衍生物可作为先导化合物。
- 抗肿瘤药物:其对多种癌细胞的抑制作用,尤其是对正常细胞的低毒性,使其成为化疗药物的潜在候选。
- 神经保护剂:结合其抗氧化活性和高BBB穿透性,稻壳酮A在神经退行性疾病的治疗中具有潜力。
- 代谢性疾病:其降血糖和抗肥胖活性值得进一步探索。
尽管前景广阔,稻壳酮A的开发仍面临诸多挑战:
1. 来源问题:天然来源含量低,化学合成步骤复杂、成本高。未来需发展高效的生物合成或半合成方法。
2. 药代动力学优化:低水溶性和潜在的快速代谢是主要瓶颈。通过结构修饰(如引入极性基团、稳定内酯环)或制剂技术可望改善。
3. 选择性提高:虽然对正常细胞毒性较低,但需进一步评估其在体内的选择性指数。
4. 机制阐明:尽管已发现多个靶点,但稻壳酮A的直接蛋白靶标尚未明确。通过化学蛋白质组学、表面等离子体共振(SPR)等技术鉴定其直接靶点,将有助于理性药物设计。
稻壳酮A作为一种源自水稻的海螺烷二萜内酯,在植物化学防御和化感作用中扮演着关键角色,同时展现出多样化的哺乳动物药理活性。其独特的化学结构、符合Lipinski规则的类药性、以及初步的安全性评价结果,使其成为一个值得深入研究的天然产物先导化合物。然而,低水溶性和有限的药代动力学数据是制约其临床转化的主要障碍。未来的研究应聚焦于:阐明其直接分子靶点、优化其药代动力学特性、发展高效可持续的制备方法,并系统评估其体内药效与安全性。通过多学科交叉合作,稻壳酮A有望从一种古老的植物防御分子,转变为现代农业和医药领域的新型工具分子。

版权所有:© 成都普瑞法科技开发有限公司(2015)备案号:蜀ICP备15035167号-1 客服热线:400-829-7929
技术支持:南京库价















