
产品名称: 四乙酰植物鞘氨醇
英文名: Tetraacetylphytosphingosine
中文别名: 四乙酰基植物鞘氨醇
英文别名: Tetraacetylphytospingosine;Phytosphingosine tetraacetate; Tetraacetyl-4-hydroxysphinganine
Cas 号: 13018-48-9
产品编码:BP2294
分子式: C26H47NO7
分子量: 485.662
来源:
化合物类型: 生物碱类(Alkaloids)
纯度: 95%~99%
分析方法: HPLC-DAD or/and HPLC-ELSD
鉴定方法: 质谱(Mass), 核磁(NMR)
包装: 棕色小玻璃瓶,按客户需求包装。
存储: 贮存在避光密闭容器中,冷藏或者冷冻长期保存。
样品溶液最好临用新配。如果需要提前配制的话,最好分成独立包装冷冻保存(-20℃以下),临用前再取出解冻,通常可以保存2周。
可以满足克级以上大量需求,详情请咨询。
四乙酰植物鞘氨醇是一种多不饱和脂肪酸。它已被证明在皮肤细胞中具有抗癌活性,可用于治疗痤疮。
四乙酰植物鞘氨醇在胺的存在下没有活性,但它被小麦胚芽中的一种叫做酪氨酸酶的酶激活。这种化合物抑制酪氨酸酶的活性,从而导致黑色素的产生减少。
四乙酰植物鞘氨醇还可以减少皮肤炎症,可以用作保湿剂。它还通过与细胞表面脂质结合的能力,对某些细菌(如结核分枝杆菌和鸟分枝杆菌复合体)的生长有抑制作用。
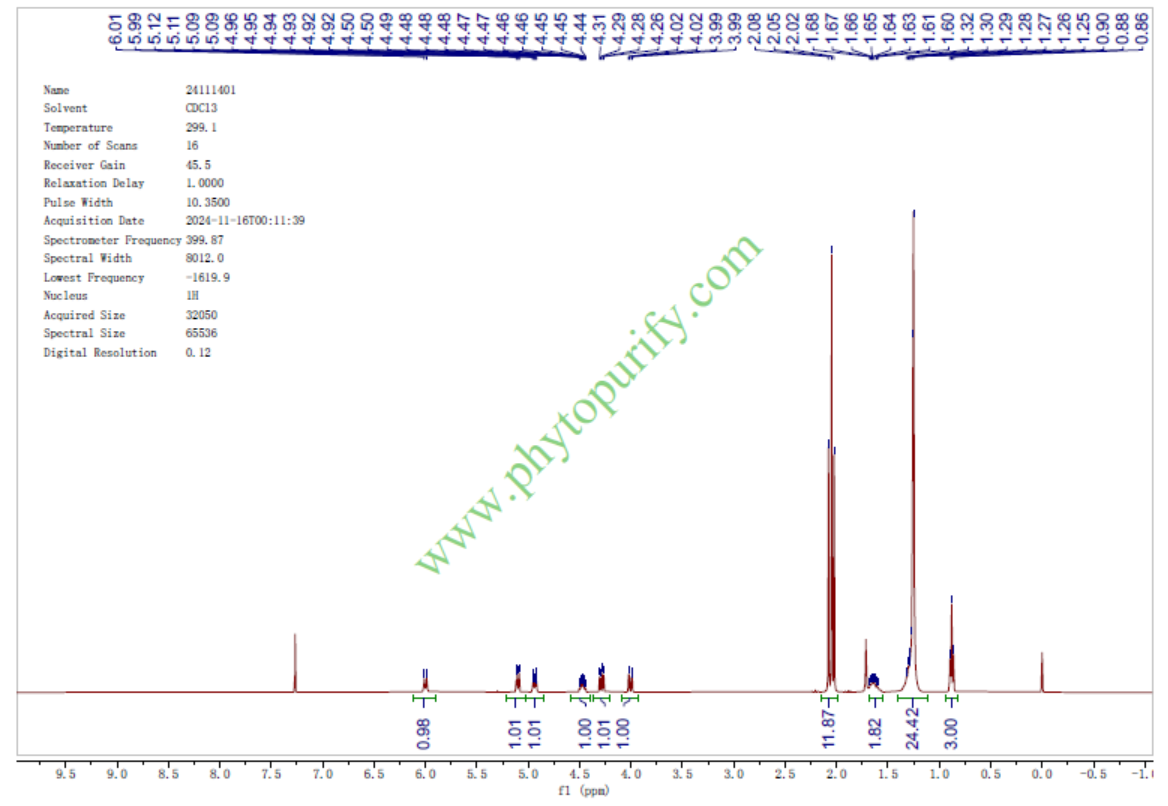
四乙酰植物鞘氨醇的核磁图谱
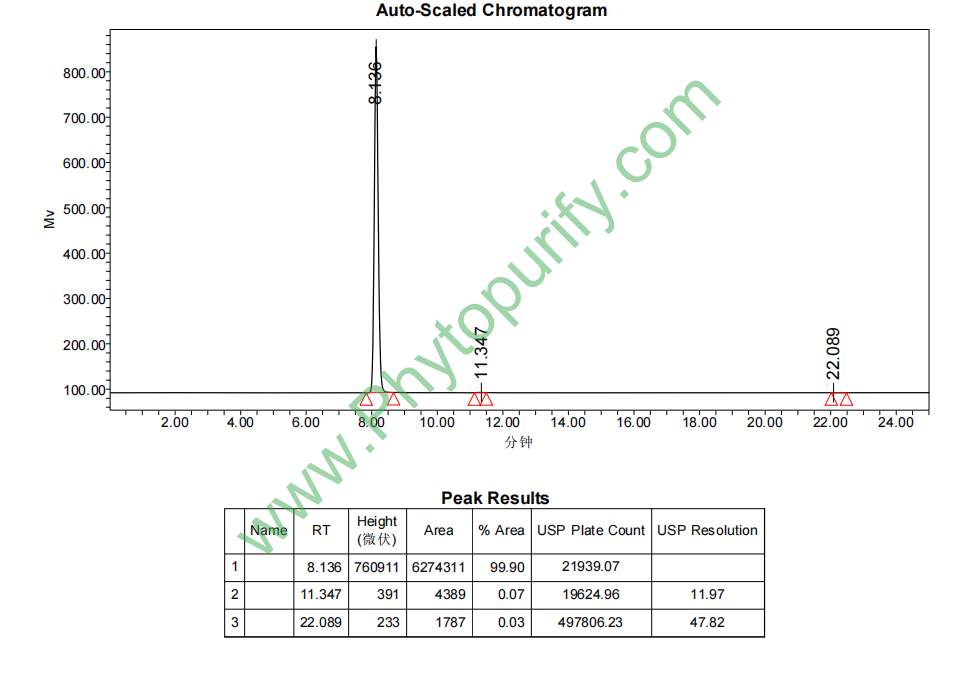
四乙酰植物鞘氨醇的HPLC图谱
储存条件:短期保存 2~8℃,长期保存 -20 ~ -80℃
108.00
4.54
4.54
0.02
0.58
0.98
低
89.33
3.57
是
否
否
否
无
无
0.0
否
否
是
是

鞘脂类化合物(Sphingolipids)是一类广泛存在于真核生物细胞膜中的结构性和功能性脂质分子。长期以来,它们被视为细胞膜的结构骨架成分。然而,近三十年的研究彻底改变了这一认知,揭示出鞘脂及其代谢产物是参与调控细胞增殖、分化、衰老、凋亡、迁移及炎症反应等多种关键生命活动的信号分子。在众多鞘脂代谢物中,植物鞘氨醇(Phytosphingosine, PHS)及其衍生物因其独特的生物学功能和潜在的治疗价值而备受关注。植物鞘氨醇最初在植物和酵母中被发现,是鞘脂代谢途径中的一个重要中间体,与哺乳动物中的鞘氨醇(Sphingosine)结构相似,但在C4位多了一个羟基。
四乙酰植物鞘氨醇(Tetraacetylphytosphingosine, TAPS),CAS号为13018-48-9,是植物鞘氨醇的四个羟基(包括C1、C3、C4位的羟基以及C2位的氨基)全部被乙酰化修饰后形成的衍生物。这种化学修饰显著改变了母体分子的理化性质和生物学活性。作为一种鞘脂代谢物,TAPS在天然产物药理学领域展现出独特的双重调控特性:一方面,它通过抑制丝裂原活化蛋白激酶(MAPK)的活化和细胞内钙离子浓度的增加,发挥显著的抗血管生成作用;另一方面,它能够诱导人永生化角质形成细胞(HaCaT细胞)发生凋亡。这些发现将TAPS与多种病理过程紧密联系起来,特别是那些涉及异常血管新生和细胞过度增殖的疾病,如肿瘤、动脉粥样硬化、银屑病和炎症性肠病等。
本文旨在对四乙酰植物鞘氨醇进行全面的专业综述,系统阐述其化学结构与理化性质、植物来源与提取方法、药理活性研究、作用机制与分子靶点、成药性评价与药代动力学特征,并展望其在临床转化中的应用前景与挑战,以期为该天然产物的深入研究与开发提供参考。
四乙酰植物鞘氨醇的化学本质是植物鞘氨醇的全乙酰化衍生物。植物鞘氨醇((2S,3S,4R)-2-氨基十八烷-1,3,4-三醇)本身是一个含有18个碳原子的长链碱基,其结构特征在于C2位有一个氨基,C1、C3、C4位各有一个羟基。TAPS通过乙酰化反应,将这四个活性基团(一个氨基和三个羟基)全部转化为乙酰基衍生物,即生成N-乙酰基和三个O-乙酰基。因此,TAPS的化学名为N-乙酰基-1,3,4-三-O-乙酰基植物鞘氨醇,其分子式为C26H47NO7。
该化合物的分子量为485.66 Da,属于中等大小的脂溶性分子。其油水分配系数(LogP)为4.541,表明其具有较高的亲脂性,这与其长脂肪链和乙酰基的疏水特性相符。高LogP值意味着TAPS在水性环境中的溶解度较差,其计算水溶性仅为0.0197 mg/mL,这对其在生物体内的吸收、分布和制剂开发提出了挑战。极性表面积(TPSA)为108.00 Ų,这一数值虽然略高于传统口服药物“规则”中通常建议的小于140 Ų的标准,但仍处于可接受的范围内,提示其具有一定的膜通透性潜力,但可能依赖于特定的转运机制或制剂技术。
从结构-活性关系(SAR)的角度来看,乙酰化修饰对TAPS的生物学功能至关重要。母体分子植物鞘氨醇本身具有强烈的细胞毒性和促凋亡活性,而乙酰化后,其毒性显著降低,同时获得了新的、更为精细的调控功能,如抗血管生成。乙酰基的引入不仅改变了分子的电荷分布和氢键供体/受体能力,还可能影响其与特定蛋白质靶点的结合模式。例如,TAPS抑制MAPK活化的能力可能与其乙酰基团介导的特定构象或疏水相互作用有关。此外,乙酰化也增强了分子的代谢稳定性,使其在体内能够更持久地发挥效应。
虽然四乙酰植物鞘氨醇在自然界中并非以高浓度游离存在,但它是植物鞘氨醇代谢途径中的一个重要修饰产物。植物鞘氨醇广泛存在于植物界,尤其是在谷物(如小麦、玉米)、大豆、酵母以及某些真菌中含量丰富。在植物中,植物鞘氨醇主要以糖基化形式(如植物鞘氨醇葡糖苷)或作为复杂鞘脂(如糖基神经酰胺)的骨架成分存在。TAPS通常是通过化学或酶学方法对天然提取的植物鞘氨醇进行乙酰化修饰而获得,而非直接从植物中大量分离。
提取TAPS或其前体植物鞘氨醇的经典流程通常包括以下几个关键步骤:
近年来,随着绿色化学理念的推广,酶法合成TAPS也受到关注。利用固定化的脂肪酶或酰基转移酶,在水-有机溶剂两相体系中,可以高效、选择性地催化植物鞘氨醇的乙酰化,避免了使用有毒的吡啶和乙酸酐。此外,超临界流体萃取(SFE)技术也被探索用于从天然原料中更高效、更环保地提取鞘脂类成分。
四乙酰植物鞘氨醇的药理活性研究主要集中在其对血管生成和细胞凋亡的调控作用,并由此衍生出对多种疾病的潜在治疗价值。
1. 抗血管生成活性
血管生成是从已有血管网络中形成新血管的过程,在肿瘤生长、转移、动脉粥样硬化斑块进展、视网膜病变和慢性炎症等多种疾病中扮演关键角色。TAPS被证实是一种有效的血管生成抑制剂。体外研究表明,TAPS能够显著抑制由血管内皮生长因子(VEGF)或碱性成纤维细胞生长因子(bFGF)诱导的人脐静脉内皮细胞(HUVECs)的增殖、迁移和管腔形成。其作用机制涉及对细胞内信号通路的干扰。具体而言,TAPS能够抑制VEGF刺激引起的MAPK(尤其是ERK1/2和p38)的磷酸化活化。MAPK通路是调控内皮细胞增殖和迁移的核心通路之一。此外,TAPS还能阻断VEGF诱导的细胞内钙离子浓度([Ca²⁺]i)的升高。钙离子作为第二信使,其浓度的动态变化对于内皮细胞激活、一氧化氮(NO)合成和细胞骨架重排至关重要。通过抑制MAPK活化和钙离子信号,TAPS有效地阻断了血管生成级联反应。体内实验,如鸡胚绒毛尿囊膜(CAM)模型和小鼠Matrigel plug实验,进一步证实了TAPS的体内抗血管生成效果,表现为新生血管密度和血红蛋白含量的显著降低。
2. 诱导细胞凋亡活性
除了抗血管生成作用,TAPS还被发现具有诱导特定类型细胞凋亡的能力。研究显示,TAPS能够以剂量和时间依赖的方式诱导HaCaT细胞(一种永生化人角质形成细胞系)发生凋亡。HaCaT细胞是研究皮肤屏障功能和银屑病等皮肤疾病的常用模型。TAPS处理后的HaCaT细胞表现出典型的凋亡形态学特征,如细胞皱缩、染色质凝聚和DNA片段化。流式细胞术分析显示,TAPS处理导致细胞周期阻滞在G0/G1期,并增加亚二倍体(Sub-G1)凋亡峰的比例。进一步的分子机制研究表明,TAPS诱导的凋亡与线粒体途径的激活有关,涉及Bcl-2家族蛋白表达的改变(如促凋亡蛋白Bax上调,抗凋亡蛋白Bcl-2下调),导致线粒体膜电位丧失,细胞色素c释放,进而激活Caspase-9和Caspase-3,最终执行凋亡程序。这种对过度增殖的角质形成细胞的选择性诱导凋亡作用,为TAPS在银屑病等增生性皮肤病中的应用提供了理论基础。
3. 抗炎与免疫调节活性
基于TAPS对MAPK和钙信号的调控,以及其与多种炎症性疾病(如动脉粥样硬化、炎症性肠病、多发性硬化)的关联,其抗炎和免疫调节活性也受到了关注。MAPK通路是炎症反应的核心驱动者,调控多种促炎细胞因子(如TNF-α、IL-6、IL-1β)的产生。通过抑制MAPK活化,TAPS可能发挥广谱的抗炎效应。此外,TAPS对细胞内钙信号的调节也可能影响免疫细胞的激活和功能,如T细胞受体信号传导和肥大细胞脱颗粒。虽然目前直接针对TAPS抗炎活性的系统研究尚不如其抗血管生成研究深入,但其靶点网络(如AMPK、TLR4、STAT3等)强烈提示其具有潜在的抗炎和免疫调节能力。例如,AMPK的激活通常与抗炎和代谢稳态相关,而TLR4是先天免疫的关键受体。
四乙酰植物鞘氨醇的药理作用是多靶点、多通路协同作用的结果。基于现有研究,其核心作用机制可归纳为以下几个方面:
1. 抑制MAPK信号通路
MAPK家族包括ERK、JNK和p38三条主要分支。TAPS被证实能够抑制VEGF等刺激因子诱导的ERK1/2和p38的磷酸化。ERK通路主要调控细胞增殖和分化,而p38通路则与应激反应、炎症和细胞迁移密切相关。TAPS可能通过直接或间接作用于MAPK通路的上游激酶(如Raf、MEK)或磷酸酶,从而阻断信号的级联放大。这一机制是其抗血管生成活性的核心。
2. 调控细胞内钙离子稳态
TAPS能够抑制激动剂诱导的细胞内钙离子浓度升高。这可能通过影响细胞膜上的钙离子通道(如瞬时受体电位通道TRPV1,该靶点与银屑病相关)或内质网上的IP3受体(如RYR2,与肿瘤血管生成相关)来实现。钙离子信号的抑制直接影响了下游的钙调蛋白依赖性激酶(CaMK)和钙调磷酸酶(Calcineurin)的活性,进而调控细胞骨架重组、NO合成和基因转录。
3. 调节细胞凋亡相关蛋白
TAPS诱导HaCaT细胞凋亡的机制涉及线粒体途径。它通过调节Bcl-2家族蛋白的平衡(上调Bax,下调Bcl-2和MCL1),增加线粒体外膜的通透性,导致细胞色素c释放,激活Caspase级联反应。此外,TAPS还可能通过影响死亡受体途径(如TNF受体)或内质网应激途径来协同诱导凋亡。其靶点列表中的MCL1和BCL2直接支持了这一机制。
4. 激活AMPK信号通路
AMPK(AMP活化蛋白激酶)是细胞能量代谢的关键传感器。靶点列表显示AMPK(PRKAA1)是TAPS在多种疾病(动脉粥样硬化、炎症性肠病、银屑病)中的共同靶点。AMPK的激活通常具有抗炎、抗增殖和促进代谢健康的作用。TAPS可能通过增加细胞内AMP/ATP比例或直接作用于AMPK的上游激酶(如LKB1)来激活AMPK。AMPK的激活可以抑制mTOR通路,从而抑制细胞生长和增殖,并可能通过磷酸化eNOS来改善血管功能,这与抗动脉粥样硬化作用相关。
5. 干预脂质代谢与炎症通路
在动脉粥样硬化背景下,TAPS的靶点包括LOX-1(凝集素样氧化低密度脂蛋白受体-1)、ABCA1(ATP结合盒转运体A1)和EHMT2。LOX-1是内皮细胞摄取氧化低密度脂蛋白(ox-LDL)的主要受体,其激活会引发内皮功能障碍和炎症。TAPS可能通过抑制LOX-1的表达或功能来减轻ox-LDL的损伤。ABCA1参与胆固醇逆向转运,其上调有助于清除血管壁中的过量胆固醇。EHMT2是一种组蛋白甲基转移酶,参与表观遗传调控。TAPS对这些靶点的调控,暗示其在改善脂质代谢和抑制动脉粥样硬化炎症方面具有潜力。
6. 调控免疫与炎症转录因子
TAPS的靶点网络还包括NFKB1(NF-κB p50亚基)、STAT3和RORC(维甲酸相关孤儿受体γ)。NF-κB和STAT3是关键的促炎转录因子,调控大量炎症和免疫相关基因的表达。RORC是Th17细胞分化的关键转录因子,在自身免疫性疾病(如银屑病、多发性硬化)中发挥核心作用。TAPS可能通过抑制这些转录因子的活性或核转位,来发挥抗炎和免疫调节作用。
评估一个天然产物能否成为候选药物,需要综合考虑其成药性(Drug-likeness)和药代动力学(ADME)特性。四乙酰植物鞘氨醇在这方面的表现呈现出机遇与挑战并存的局面。
1. 成药性评价
根据Lipinski的“五规则”(Rule of Five),TAPS的分子量(485.66 Da)略高于500 Da的阈值,LogP值(4.541)也略高于5的阈值,这提示其可能存在口服生物利用度不佳的问题。其TPSA(108.00 Ų)和氢键供体/受体数目(符合规则)则是有利因素。综合来看,TAPS是一个处于“规则边缘”的分子,其口服吸收可能受限,但并非绝对不可能,需要借助制剂手段来改善。
2. 药代动力学特征(预测与推断)
3. 安全性评价
基于四乙酰植物鞘氨醇独特的药理活性和多靶点作用机制,其在以下疾病领域的临床应用前景值得深入探索:
1. 肿瘤治疗与抗血管生成疗法
TAPS最明确的应用前景在于其抗血管生成活性。肿瘤的生长和转移高度依赖新生血管。作为一种新型的血管生成抑制剂,TAPS可以单独使用或与化疗、放疗及其他靶向药物(如VEGF抑制剂)联合使用,以增强抗肿瘤效果并克服耐药性。其独特的机制(同时抑制MAPK和钙信号)可能为现有抗血管生成治疗提供一种补充或替代方案。特别是对于某些对VEGF抑制剂不敏感的肿瘤类型,TAPS可能具有优势。
2. 银屑病等增生性皮肤病
TAPS能够诱导角质形成细胞(HaCaT)凋亡,且其靶点包括银屑病发病机制中的关键分子(如STAT3、RORC、AMPK)。这使其成为治疗银屑病的有希望的候选药物。与目前常用的糖皮质激素或维生素D3衍生物相比,TAPS可能提供一种新的作用机制,尤其适用于对传统治疗无效或产生耐药的患者。局部外用制剂(如软膏、乳膏)是其最直接的开发形式,可以最大限度地发挥局部药效,同时减少全身性副作用。
3. 动脉粥样硬化与心血管疾病
TAPS通过抑制LOX-1、激活AMPK、调节胆固醇转运(ABCA1)和抗炎等多重机制,有望用于防治动脉粥样硬化。它可以抑制血管内皮细胞的激活和泡沫细胞的形成,稳定动脉粥样硬化斑块,从而预防心肌梗死和脑卒中等心血管事件。然而,如何实现有效的口服吸收和靶向血管壁的递送,是开发其心血管适应症的主要挑战。
4. 炎症性肠病与自身免疫性疾病
TAPS对MAPK、NF-κB、STAT3和TLR4等炎症信号通路的抑制作用,使其在治疗炎症性肠病(如克罗恩病、溃疡性结肠炎)和多发性硬化等自身免疫性疾病中具有潜力。通过口服或直肠给药(针对IBD),TAPS可能局部抑制肠道炎症。对于多发性硬化,由于其BBB通透性低,可能需要开发特殊的脑靶向递送系统(如纳米粒、脂质体)或利用其作为前体药物的特性,使其在体内代谢后产生能够进入中枢的活性成分。
未来展望与挑战:
四乙酰植物鞘氨醇作为一种源自天然鞘脂代谢的独特分子,通过其乙酰化修饰实现了从细胞毒性分子向多功能信号调控因子的华丽转身。它集抗血管生成、诱导特定细胞凋亡、抗炎和潜在免疫调节活性于一身,其作用机制涉及MAPK、钙信号、AMPK、Bcl-2家族以及NF-κB、STAT3等多个关键信号通路和靶点,展现出多靶点、网络化调控的复杂药理学特征。这一特性使其在肿瘤、银屑病、动脉粥样硬化和自身免疫性疾病等多个重大疾病领域展现出诱人的治疗潜力。
然而,从实验室发现到临床应用的道路依然充满挑战。其固有的理化性质(高亲脂性、低水溶性)所带来的药代动力学难题是首要障碍。未来的研究重点应聚焦于创新制剂技术的开发、深入的代谢与药代动力学表征、关键靶点的精确验证以及系统的体内药效与安全性评价。随着对这些问题的逐步攻克,四乙酰植物鞘氨醇及其衍生物有望成为一类具有独特价值的新型治疗药物,为人类健康事业做出贡献。对这类“边缘”天然产物的深入研究,也为我们理解鞘脂信号网络的复杂性以及开发基于天然产物的创新药物提供了宝贵的范例。

版权所有:© 成都普瑞法科技开发有限公司(2015)备案号:蜀ICP备15035167号-1 客服热线:400-829-7929
技术支持:南京库价















