
产品名称: 乳糖-N-新四糖
英文名: Lacto-N-neotetraose
中文别名:
英文别名: N-Neotetraose;Lacto-N-neo-tetraose; Lacto-N-neotetraose (LNnT); LNnT
Cas 号: 13007-32-4
产品编码:BP5436
分子式: C26H45NO21
分子量: 707.632
来源:
化合物类型: 寡糖(Oligoses)
纯度: 95%~99%
分析方法: HPLC-DAD or/and HPLC-ELSD
鉴定方法: 质谱(Mass), 核磁(NMR)
包装: 棕色小玻璃瓶,按客户需求包装。
存储: 贮存在避光密闭容器中,冷藏或者冷冻长期保存。
样品溶液最好临用新配。如果需要提前配制的话,最好分成独立包装冷冻保存(-20℃以下),临用前再取出解冻,通常可以保存2周。
可以满足克级以上大量需求,详情请咨询。
提供相关图谱和分析方法指导,可以提供包括色谱柱,标准品和分析方法在内的含量测定整体服务包,价格优惠。
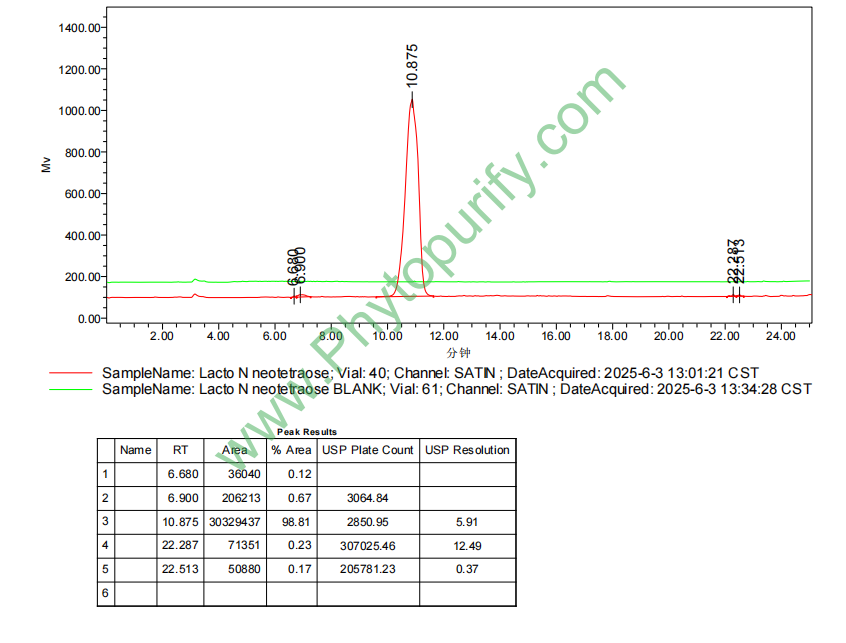
乳糖-N-新四糖的HPLC图谱
储存条件:短期保存 2~8℃,长期保存 -20 ~ -80℃
396.11
-8.00
-8.00
否
10.00
否
否
否
否
否

天然产物作为药物发现的重要源泉,在人类健康维护与疾病治疗中扮演着不可替代的角色。在众多天然来源的活性分子中,母乳低聚糖(Human Milk Oligosaccharides, HMOs)因其独特的结构多样性与生物学功能,近年来成为糖生物学与营养药理学领域的研究热点。乳糖-N-新四糖(Neolactotetraose,简称LNnT)作为HMOs中含量较为丰富且结构特征鲜明的成员之一,其生物学意义与药理学潜力正逐步被揭示。
乳糖-N-新四糖,化学名称为β-D-半乳糖-(1→4)-N-乙酰基-β-D-葡萄糖胺-(1→3)-β-D-半乳糖-(1→4)-D-葡萄糖,是一种由四个单糖单元组成的线性四糖。其CAS登记号为13007-32-4,分子式为C26H45NO21。作为母乳中天然存在的低聚糖成分,LNnT在婴儿肠道微生态建立、免疫系统发育以及抵御病原微生物感染等方面发挥着关键作用。与另一结构异构体乳糖-N-四糖(Lacto-N-tetraose, LNT)相比,LNnT的糖苷键连接方式不同,导致其空间构象与生物学活性存在显著差异。
近年来,随着合成生物学与酶工程技术的进步,LNnT的规模化制备成为可能,这极大地推动了其作为功能性食品成分或潜在治疗药物的研究进程。从天然产物药理学视角审视,LNnT不仅是一种营养因子,更是一种具有明确分子靶点与信号通路调控作用的生物活性分子。本文旨在系统梳理LNnT的化学结构特征、天然来源、提取制备方法、药理活性、作用机制、成药性评价及临床应用前景,以期为该天然产物的深入开发与转化研究提供全面的学术参考。
乳糖-N-新四糖的结构核心由四个单糖残基通过特定的糖苷键连接而成。其精确结构序列为:Galβ1-4GlcNAcβ1-3Galβ1-4Glc。从非还原端到还原端依次为:D-半乳糖(Gal)通过β-1,4糖苷键连接至N-乙酰基-D-葡萄糖胺(GlcNAc),后者再通过β-1,3糖苷键连接至第二个D-半乳糖,最终通过β-1,4糖苷键连接至末端的D-葡萄糖(Glc)。这种线性排列方式赋予了LNnT独特的分子识别特性。
与LNT(Galβ1-3GlcNAcβ1-3Galβ1-4Glc)相比,LNnT的关键差异在于非还原端半乳糖与N-乙酰氨基葡萄糖之间的连接方式:LNnT为β-1,4连接,而LNT为β-1,3连接。这一看似微小的结构差异,却导致两者在分子构象、电荷分布以及受体识别能力上的显著不同。核磁共振(NMR)与分子动力学模拟研究显示,LNnT的糖苷键柔性使其能够呈现更为伸展的构象,有利于与凝集素类受体蛋白的结合。
基于计算化学与实验测定,LNnT的关键理化参数如下:分子量为739.63 Da,属于中等大小的寡糖分子。其脂水分配系数(LogP)极低,为-8.0,表明该化合物具有极强的亲水性,几乎不溶于有机溶剂,而极易溶于水。拓扑极性表面积(TPSA)高达396.11 Ų,这与其分子中大量羟基和酰胺基团的存在相一致。氢键受体数量为22个,进一步印证了其形成强氢键网络的能力。
这些理化性质决定了LNnT在生物体内的行为特征:高水溶性使其在肠道环境中能够保持溶解状态,有利于与肠道上皮细胞表面的受体相互作用;而极低的脂溶性则意味着其难以被动扩散通过细胞膜,口服给药后主要停留在胃肠道腔道内,系统吸收极为有限。此外,LNnT在酸性条件下(如胃液)相对稳定,但在某些肠道细菌产生的糖苷酶作用下可被逐步降解为单糖或二糖片段。
严格意义上,乳糖-N-新四糖并非植物来源的天然产物,而是哺乳动物乳汁中特有的低聚糖成分。在人类母乳中,LNnT是含量最为丰富的HMOs之一,尤其在初乳阶段浓度较高。研究表明,不同遗传背景的哺乳期女性乳汁中LNnT含量存在显著差异,这与个体中岩藻糖基转移酶(FUT2和FUT3)的基因多态性密切相关。除人类外,少数哺乳动物(如某些灵长类动物)的乳汁中也检测到LNnT的存在,但含量远低于人乳。
值得注意的是,LNnT在植物界中尚未被报道为天然存在的成分。然而,某些微生物(如特定乳酸菌菌株)在特定培养条件下能够合成结构类似的低聚糖,这为LNnT的微生物发酵生产提供了生物学基础。
由于LNnT在天然乳汁中含量有限且与其他HMOs共存,直接从人乳中提取LNnT既不经济也不符合伦理规范。因此,目前LNnT的获取主要依赖化学合成、酶法合成以及微生物发酵三种途径。
化学合成法:采用经典的糖化学合成策略,通过保护基操作、糖苷化反应及脱保护步骤,从单糖或二糖单元逐步构建LNnT的四糖骨架。该方法虽然能够获得结构明确的产物,但步骤繁琐、总收率低、需要使用大量有机溶剂和有毒试剂,不适合大规模生产。
酶法合成:利用糖基转移酶(如β-1,4-半乳糖基转移酶、β-1,3-N-乙酰氨基葡萄糖转移酶等)的底物特异性与区域选择性,以乳糖或N-乙酰氨基乳糖为受体,以核苷酸糖(如UDP-半乳糖、UDP-N-乙酰氨基葡萄糖)为供体,在体外构建LNnT。该方法反应条件温和、立体选择性高,但需要昂贵的核苷酸糖供体,且酶的成本与稳定性限制了其工业化应用。
微生物发酵法:近年来,通过代谢工程改造的大肠杆菌或酵母菌株生产LNnT成为最具前景的路线。研究者将编码糖基转移酶、糖核苷酸合成酶及糖转运蛋白的基因导入宿主菌,构建完整的LNnT生物合成途径。以乳糖为底物,通过发酵过程实现LNnT的胞内或胞外积累。发酵液经离心去除菌体后,采用活性炭吸附、离子交换色谱、凝胶过滤色谱及制备型高效液相色谱(HPLC)等步骤进行纯化,最终获得高纯度(>95%)的LNnT产品。该方法的优势在于成本相对较低、可规模化放大,且符合绿色化学理念。
LNnT最受关注的药理活性之一是其作为益生元对肠道菌群的调节作用。体外发酵实验表明,LNnT能够选择性促进双歧杆菌(尤其是长双歧杆菌婴儿亚种)和乳杆菌等有益菌的生长,同时抑制产气荚膜梭菌、大肠杆菌等潜在致病菌的增殖。这种选择性利用能力源于有益菌编码的特定糖苷水解酶(如LNnT特异性乳糖-N-生物苷酶),能够将LNnT水解为可代谢的单糖或二糖片段。
动物模型研究进一步证实,口服LNnT可显著增加小鼠盲肠内容物中双歧杆菌的相对丰度,同时降低脱硫弧菌属等促炎菌群的比例。代谢组学分析显示,LNnT干预后肠道短链脂肪酸(尤其是乙酸和丁酸)浓度升高,这与有益菌代谢LNnT产生的终产物一致。
LNnT作为可溶性糖类受体类似物,能够竞争性抑制多种病原微生物对宿主上皮细胞的黏附。机制上,LNnT的末端半乳糖-β-1,4-N-乙酰氨基葡萄糖结构域模拟了宿主细胞表面糖蛋白或糖脂上的受体表位,从而“诱捕”病原菌的黏附素(如菌毛凝集素)。
针对肠道病原菌的研究显示,LNnT可显著抑制产肠毒素大肠杆菌(ETEC)K88菌株对猪肠道上皮细胞的黏附,抑制率可达60%以上。此外,LNnT对空肠弯曲杆菌、沙门氏菌及轮状病毒的黏附也具有阻断作用。值得注意的是,LNnT对呼吸道合胞病毒(RSV)的体外抗病毒活性也被报道,提示其可能具有超越肠道的全身性抗感染潜力。
LNnT的免疫调节作用是其药理活性的另一重要维度。体外实验表明,LNnT能够直接与肠道上皮细胞表面的C型凝集素受体(如DC-SIGN、甘露糖受体)相互作用,调节下游信号通路。在树突状细胞模型中,LNnT处理可诱导IL-10、TGF-β等抗炎细胞因子的分泌,同时抑制TNF-α、IL-12等促炎因子的产生,呈现典型的抗炎免疫表型。
在肠道炎症动物模型中,口服LNnT可减轻葡聚糖硫酸钠(DSS)诱导的结肠炎症状,表现为体重下降减轻、疾病活动指数降低及结肠组织病理损伤改善。机制上,LNnT通过促进调节性T细胞(Treg)的分化与扩增,恢复肠道免疫稳态。此外,LNnT还可增强肠道屏障功能,通过上调紧密连接蛋白(如occludin、claudin-1)的表达,降低肠道通透性,减少内毒素向循环系统的易位。
新兴研究提示,LNnT可能对早期神经发育具有积极影响。在啮齿动物模型中,围产期补充LNnT可促进仔鼠海马区突触可塑性相关蛋白(如BDNF、PSD-95)的表达,改善空间学习与记忆能力。这一效应被认为与LNnT调节肠道菌群-肠-脑轴有关:通过改变肠道菌群组成,影响神经活性代谢物(如5-羟色胺前体、短链脂肪酸)的产生,进而通过迷走神经或体液途径影响中枢神经系统功能。
LNnT的生物学效应部分源于其与特定受体蛋白的直接结合。表面等离子体共振(SPR)与等温滴定量热(ITC)实验已鉴定出多个LNnT的蛋白靶点:
DC-SIGN(树突状细胞特异性ICAM-3捕获非整合素):LNnT的末端LacNAc(Galβ1-4GlcNAc)结构域能够以微摩尔级别的亲和力结合DC-SIGN的碳水化合物识别结构域(CRD)。这种结合可调控树突状细胞的成熟状态与细胞因子分泌谱。
半乳糖凝集素(Galectins):LNnT与半乳糖凝集素-1(Gal-1)和半乳糖凝集素-3(Gal-3)具有中等亲和力。Gal-3在炎症反应中发挥关键作用,LNnT对其结合可抑制Gal-3介导的细胞黏附与信号转导。
Toll样受体(TLRs):虽然LNnT并非TLR的直接激动剂或拮抗剂,但其可通过调节TLR4与TLR2的共受体(如MD-2、CD14)的膜定位,间接影响TLR信号通路的激活阈值。
LNnT对细胞内信号通路的调控呈现多靶点、网络化的特征:
NF-κB通路:在肠道上皮细胞中,LNnT可抑制TNF-α或LPS诱导的IκBα磷酸化与降解,从而阻断NF-κB的核转位,减少促炎基因(如IL-8、MCP-1)的转录。
MAPK通路:LNnT能够选择性激活p38 MAPK通路,同时抑制JNK的磷酸化,这种差异调控有助于维持细胞应激反应与存活信号的平衡。
PI3K/Akt通路:在肠上皮细胞损伤模型中,LNnT通过激活PI3K/Akt信号轴,促进细胞存活与增殖,加速受损黏膜的修复。
Wnt/β-catenin通路:LNnT可轻度激活肠道干细胞标志物Lgr5的表达,提示其可能通过Wnt信号通路参与肠道上皮的更新与再生。
除直接受体结合外,LNnT的许多药理效应是通过调节肠道菌群间接实现的。LNnT作为益生元,促进特定有益菌(如双歧杆菌)的生长,这些菌群产生的代谢产物(短链脂肪酸、色氨酸代谢物、胆汁酸衍生物)可作为信号分子作用于宿主细胞。例如,丁酸可通过抑制组蛋白去乙酰化酶(HDAC)活性,调节宿主基因表达;而吲哚-3-丙酸等色氨酸代谢物则可激活芳烃受体(AhR),增强肠道屏障功能与免疫耐受。
基于前述理化性质与计算药理学参数,LNnT的成药性特征可归纳如下:
口服生物利用度:预测值为10%,但这一数值可能高估了实际情况。由于LNnT的强亲水性与大分子量,其通过肠道上皮细胞被动扩散的能力极低。实际口服后,LNnT主要以原型形式存在于胃肠道腔道内,系统吸收率通常低于1%。
血脑屏障穿透性:预测结果为“No”,这与LNnT的极性特征一致。除非在病理状态下血脑屏障完整性受损,否则LNnT难以进入中枢神经系统。
肝毒性:预测为“No”。LNnT作为天然食物成分,在常规剂量下未观察到肝细胞毒性。但高剂量静脉给药的安全性数据尚缺乏。
心脏毒性:预测为“No”。LNnT对hERG钾通道无抑制活性,QT间期延长风险极低。
遗传毒性:Ames试验结果未知。考虑到LNnT为天然糖类分子,且无已知的致突变结构警示,其遗传毒性风险预计较低。
由于LNnT的口服系统暴露极低,传统的药代动力学参数(如Cmax、AUC、t1/2)在口服给药后难以准确测定。LNnT的药代动力学行为更适宜从“局部药代动力学”角度理解:
吸收:口服后,LNnT在胃和小肠中几乎不被吸收。少量可能通过细胞旁路途径或受体介导的转胞吞作用进入门静脉循环,但比例极低。
分布:系统吸收的LNnT主要分布于血液中,由于分子量大且亲水,难以穿透毛细血管壁进入组织间隙。
代谢:LNnT在人体内不被宿主酶系代谢。其降解主要依赖肠道菌群编码的糖苷水解酶,降解产物(半乳糖、N-乙酰氨基葡萄糖、乳糖)可被宿主吸收利用。
排泄:未被吸收的LNnT随粪便排出体外。系统吸收的部分可能通过肾脏以原型形式排泄,但这一途径的贡献极小。
LNnT作为母乳中天然存在的成分,具有较高的安全性阈值。在动物毒理学研究中,大鼠口服LNnT的NOAEL(未观察到有害作用的剂量)超过5 g/kg体重/天。人体临床研究显示,健康成人每日摄入10-20克LNnT未报告严重不良事件,仅少数受试者出现轻度腹胀或排气增多。婴儿配方奶粉中添加LNnT(浓度接近母乳水平)已被多个国家的监管机构批准,进一步证实了其安全性。
LNnT最直接的临床应用方向是作为婴儿配方奶粉的功能性强化成分。母乳喂养婴儿的肠道菌群以双歧杆菌为主导,而配方奶粉喂养婴儿的菌群多样性较低且潜在致病菌比例较高。在配方奶粉中添加LNnT可部分模拟母乳的低聚糖组成,促进双歧杆菌定植,降低坏死性小肠结肠炎(NEC)和感染性腹泻的风险。目前,欧洲食品安全局(EFSA)已批准LNnT作为新型食品成分用于婴儿配方食品。
基于其抗炎与屏障保护作用,LNnT在炎症性肠病(IBD)治疗中具有潜在应用价值。临床前研究已证实LNnT对DSS结肠炎模型的保护作用,未来需开展随机对照临床试验验证其在溃疡性结肠炎或克罗恩病患者中的疗效。此外,LNnT作为益生元与现有IBD治疗药物(如美沙拉嗪、生物制剂)联用,可能产生协同效应。
LNnT的抗黏附特性使其成为预防肠道感染的候选药物。针对旅行者腹泻、抗生素相关性腹泻以及医院获得性艰难梭菌感染,LNnT可作为非抗生素的预防策略。此外,LNnT对呼吸道病毒(如RSV)的抑制活性提示其可能用于呼吸道感染的预防,但需解决局部给药(如鼻喷雾剂)的制剂问题。
新兴研究提示,LNnT可能通过调节肠道菌群-免疫轴,对肥胖、2型糖尿病及非酒精性脂肪性肝病(NAFLD)等代谢性疾病产生有益影响。LNnT促进的丁酸产生可改善胰岛素敏感性,而抗炎作用可减轻脂肪组织炎症。此外,LNnT在食物过敏预防中的潜力也值得探索,其免疫调节特性可能有助于诱导口服耐受。
尽管LNnT的临床应用前景广阔,但仍面临若干挑战:首先,大规模生产成本仍需进一步降低,以满足食品与药品领域的经济性要求;其次,LNnT在体内的代谢命运与生物活性片段尚需深入研究,以阐明其构效关系;第三,针对特定适应症的临床证据链尚不完整,需要更多高质量的人体研究;最后,LNnT与其他HMOs(如2′-岩藻糖基乳糖、LNT)的协同效应值得系统探索,以开发更接近母乳功能的复合配方。
乳糖-N-新四糖作为母乳低聚糖家族的重要成员,其天然产物药理学价值正随着研究的深入而日益彰显。从化学结构角度看,LNnT的线性四糖骨架与特定的糖苷键连接方式赋予了其独特的分子识别能力;从药理活性角度看,LNnT通过直接受体结合与间接菌群调节的双重机制,发挥益生元、抗感染、免疫调节及肠道屏障保护等多重生物学效应;从成药性角度看,LNnT具有优异的安全性特征,但口服生物利用度低的特点决定了其适宜作为局部作用(肠道)的药物或功能性食品成分。
展望未来,随着合成生物学技术的进步与临床转化研究的推进,LNnT有望从母乳中的天然成分发展为预防和治疗肠道疾病、免疫紊乱及代谢综合征的新型药物或营养制剂。这一过程不仅需要糖化学、微生物学、免疫学与药理学的跨学科协作,更需要学术界与产业界的紧密合作,以推动这一天然产物从实验室走向临床应用,最终造福人类健康。

版权所有:© 成都普瑞法科技开发有限公司(2015)备案号:蜀ICP备15035167号-1 客服热线:400-829-7929
技术支持:南京库价















