
产品名称: N-乙酰-神经氨酸(燕窝酸)
英文名: N-acetylneuraminic acid
中文别名: 唾液酸;燕窝酸
英文别名:
Cas 号: 131-48-6
产品编码:BP4175
分子式: C11H19NO9
分子量: 309.271
来源:
物理性状:
化合物类型: 脂肪族类(Aliphatics)
纯度: 95%~99%
分析方法: HPLC-DAD or/and HPLC-ELSD
鉴定方法: 质谱(Mass), 核磁(NMR)
包装: 棕色小玻璃瓶,按客户需求包装。
存储: 贮存在避光密闭容器中,冷藏或者冷冻长期保存。
样品溶液最好临用新配。如果需要提前配制的话,最好分成独立包装冷冻保存(-20℃以下),临用前再取出解冻,通常可以保存2周。
可以满足克级以上大量需求,详情请咨询。
提供相关图谱和分析方法指导,可以提供包括色谱柱,标准品和分析方法在内的含量测定整体服务包,价格优惠。
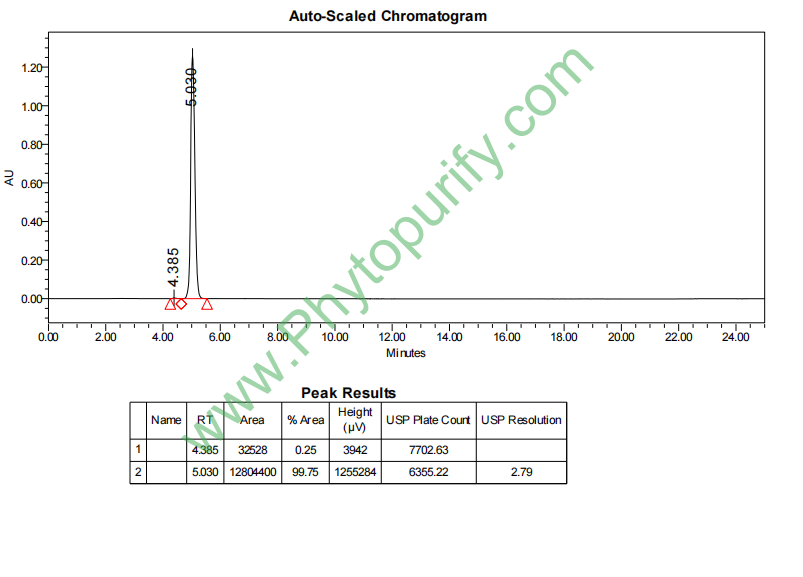
N-乙酰-神经氨酸的HPLC图谱
储存条件:短期保存 2~8℃,长期保存 -20 ~ -80℃
176.78
-2.58
-4.15
161.60
0.44
0.20
低
26.98
4.21
是
否
否
否
无
无
0.0
是
是
是
是

在生命科学的广阔图景中,糖类分子,尤其是唾液酸,正逐渐从简单的能量储存单元和结构组分,转变为介导复杂生物过程的关键信息分子。唾液酸是一类九碳酮糖酸的衍生物,广泛存在于脊椎动物细胞表面糖蛋白和糖脂的末端。其中,N-乙酰-神经氨酸(N-acetylneuraminic acid,Neu5Ac)是自然界中分布最广、含量最丰富的一种唾液酸,也是众多唾液酸家族成员的生物合成前体。因其最初从燕窝中分离并得到高浓度富集,故在商业和大众语境中常被称为“燕窝酸”。
Neu5Ac的发现历史可追溯至20世纪中叶。1936年,Gunnar Blix从牛下颌黏蛋白中分离出一种酸性糖类物质;几乎同时,Ernst Klenk从脑苷脂中分离出“神经氨酸”。后续研究证实二者为同一类物质,并最终确定了其化学结构。随后的数十年里,科学家们逐步揭示了Neu5Ac在细胞识别、信号转导、病原体感染、免疫调节及神经发育等过程中的核心作用。特别是在免疫学领域,Neu5Ac作为细胞表面糖链末端的“哨兵”,通过与多种免疫受体(如Siglecs、选择素等)的相互作用,精细调控着先天性和适应性免疫应答的平衡。
近年来,随着糖生物学和免疫药理学的交叉融合,对Neu5Ac的研究已从基础的结构与功能描述,深入至其在疾病干预中的潜在应用价值。其作为免疫调节剂的潜力尤为引人注目,涉及从抗炎、抗病毒到抗肿瘤免疫的广阔领域。本文旨在系统综述N-乙酰-神经氨酸的化学特性、来源、药理活性、作用机制及成药性前景,以期为该天然产物的深入研究和临床转化提供全面的学术参考。
N-乙酰-神经氨酸的化学本质为5-乙酰氨基-3,5-二脱氧-D-甘油基-D-半乳糖-2-酮糖酸。其核心骨架是一个九碳酮糖酸,具有独特的吡喃糖环结构。在生理pH条件下,其C2位的羧基(pKa约2.6)主要以解离的羧酸根形式存在,赋予分子负电荷,这是其参与静电相互作用和离子交换的关键。C5位上的乙酰氨基(-NHCOCH₃)是其区别于其他唾液酸(如N-羟乙酰神经氨酸,Neu5Gc)的特征基团。
从立体化学角度看,Neu5Ac存在α和β两种异头构型。在天然糖缀合物中,唾液酸几乎总是以α-糖苷键(α2-3、α2-6或α2-8)连接于半乳糖(Gal)、N-乙酰半乳糖胺(GalNAc)或其他唾液酸残基上。然而,游离的Neu5Ac在溶液中主要以β-异头体形式存在,即N-乙酰基-β-神经氨酸。该β构型是α构型的前体,也是许多唾液酸转移酶和唾液酸酶识别与作用的底物。作为N-乙酰基-β-神经氨酸酯的共轭酸,其在体内外扮演着“表位”的角色,即作为被免疫系统或特定受体识别的最小结构单元。
在理化性质方面,Neu5Ac的分子量为309.27 g/mol,计算所得油水分配系数(LogP)为-2.58,表明其具有极强的亲水性,几乎不溶于非极性有机溶剂。其极性表面积(TPSA)高达176.78 Ų,主要由羧基、羟基和乙酰氨基贡献,进一步证实了其高水溶性(计算水溶性为161.6 mg/mL)。这些特性决定了其口服生物利用度可能受限,且不易穿透血脑屏障(BBB渗透性低)。此外,基于计算机模拟的成药性评估显示,Neu5Ac对hERG钾通道无抑制风险(hERG抑制:否),并且在Ames试验中呈阴性(Ames试验:0.0),提示其遗传毒性风险极低,为后续的药物开发提供了良好的安全性基础。
尽管唾液酸在动物界中广泛存在,但在植物界中的含量通常极低甚至难以检测。因此,严格来说,Neu5Ac并非典型的“植物来源”天然产物。其最著名的天然富集来源是金丝燕(属雨燕科)的唾液腺分泌物——燕窝。燕窝中唾液酸含量高达7%-12%(干重),是已知天然生物材料中含量最高的。此外,哺乳动物的乳汁(尤其是初乳)、鸡蛋、以及某些动物组织(如脑、颌下腺)也是Neu5Ac的重要来源。
由于直接从天然原料中提取成本高昂且产量有限,目前工业上生产Neu5Ac主要依赖生物技术方法,包括:
1. 酶法合成:利用N-乙酰神经氨酸醛缩酶(Neu5Ac aldolase),以N-乙酰-D-甘露糖胺(ManNAc)和丙酮酸为底物,通过逆醛缩反应高效合成Neu5Ac。该方法具有反应条件温和、立体选择性高、副产物少的优点,是目前的主流生产工艺。
2. 微生物发酵法:通过基因工程改造大肠杆菌或枯草芽孢杆菌等微生物,使其过量表达合成Neu5Ac的关键酶(如GlcNAc 2-epimerase和Neu5Ac aldolase),并以廉价碳源(如葡萄糖、甘油)为底物进行发酵生产。该方法成本更低,适合大规模工业化生产。
3. 化学合成法:以糖类衍生物为起始原料,通过多步化学反应合成Neu5Ac。该方法步骤繁琐、产率低且涉及有毒试剂,主要用于实验室研究或同位素标记化合物的制备。
从天然原料中提取Neu5Ac的经典流程包括:原料(如燕窝或蛋清)经水或稀酸提取,通过离心、过滤去除不溶性杂质,然后利用离子交换层析(如阴离子交换树脂)进行分离纯化,最后经脱盐、浓缩和冷冻干燥获得高纯度产品。现代工艺常结合膜分离技术(超滤、纳滤)和活性炭脱色,以进一步提高纯度和效率。
N-乙酰-神经氨酸的药理活性研究已从早期的营养学观察,拓展至多个疾病领域的分子药理学层面,尤其在免疫调节、抗病毒和神经保护方面展现出显著潜力。
1. 免疫调节活性
这是Neu5Ac最受关注的核心药理活性。大量研究表明,外源性补充Neu5Ac能够显著调节机体免疫功能。
* 增强体液免疫:在动物模型和人体临床试验中,Neu5Ac(特别是燕窝提取物)被发现能促进脾脏淋巴细胞增殖,提高血清中免疫球蛋白(如IgG、IgM)水平,并增强疫苗(如流感疫苗)的抗体应答。其机制可能与激活B细胞和辅助性T细胞有关。
* 调节细胞免疫:Neu5Ac对T细胞亚群的分化具有双向调节作用。一方面,它可通过促进Th1型细胞因子(如IFN-γ)的分泌,增强细胞毒性T淋巴细胞(CTL)的活性,从而提升抗病毒和抗肿瘤免疫。另一方面,它又能诱导调节性T细胞(Treg)的分化,上调抗炎因子IL-10和TGF-β的表达,抑制过度炎症反应,显示出在自身免疫性疾病和炎症性疾病中的潜在治疗价值。
* 抗炎活性:在脂多糖(LPS)诱导的巨噬细胞炎症模型中,Neu5Ac能够显著抑制促炎因子(如TNF-α、IL-6、IL-1β)的产生,同时上调抗炎因子IL-10的水平。这种抗炎作用与其对NF-κB和STAT3信号通路的调控密切相关。
2. 抗病毒活性
Neu5Ac是许多病毒(如流感病毒、副流感病毒、轮状病毒)入侵宿主细胞的“门户”。病毒表面的血凝素(HA)蛋白特异性识别宿主细胞表面的唾液酸(主要是α2-6或α2-3连接的Neu5Ac),从而介导病毒吸附和膜融合。因此,外源性的游离Neu5Ac或含Neu5Ac的糖缀合物可作为“诱饵受体”,竞争性地与病毒HA结合,从而阻断病毒对宿主细胞的感染。这一原理已被成功应用于抗流感药物扎那米韦(Zanamivir)和奥司他韦(Oseltamivir)的设计中,它们作为唾液酸类似物,通过抑制神经氨酸酶(NA)的活性,阻止病毒从感染细胞中释放和扩散。
3. 神经保护与认知改善
作为大脑中含量最丰富的唾液酸,Neu5Ac是神经节苷脂(Gangliosides)的重要组成部分,参与神经元的生长、分化、突触形成和可塑性调节。动物实验表明,孕期和哺乳期补充Neu5Ac可显著提高子代大鼠的学习记忆能力。在阿尔茨海默病(AD)模型中,Neu5Ac被发现能减少β-淀粉样蛋白(Aβ)的聚集和神经毒性,并改善认知功能。其机制可能涉及调节神经递质释放、抑制氧化应激和神经炎症。
4. 抗肿瘤活性
Neu5Ac在肿瘤免疫中的角色具有双重性。一方面,肿瘤细胞表面过度表达的唾液酸(如α2-6连接的Neu5Ac)可与免疫细胞(如NK细胞、巨噬细胞)表面的抑制性受体Siglec-7和Siglec-9结合,传递“别吃我”信号,从而逃避免疫监视。另一方面,某些研究也发现,特定形式的Neu5Ac衍生物或通过调节肿瘤微环境中的免疫细胞功能,可能发挥抗肿瘤作用。例如,通过抑制肿瘤相关巨噬细胞(TAMs)的M2型极化,增强抗肿瘤免疫应答。
Neu5Ac的多种药理活性根植于其与特定分子靶点的精细相互作用。在免疫调节领域,其作用网络尤为复杂,涉及多个关键信号通路和转录因子。
1. 模式识别受体(PRRs)的调控
* TLR4(Toll样受体4):TLR4是识别LPS等病原相关分子模式(PAMPs)的关键受体,启动先天免疫反应。研究发现,Neu5Ac能够直接与TLR4/MD2复合物结合,其作用具有浓度依赖性。在低浓度下,它可能作为TLR4的弱激动剂,适度激活下游信号;而在高浓度下,它则可能作为拮抗剂,竞争性抑制LPS与TLR4的结合,从而抑制NF-κB的过度激活,发挥抗炎作用。这种“双相调节”模式是理解其免疫调节精细性的关键。
2. 信号转导与转录因子
* NF-κB(核因子κB):NF-κB是炎症反应的核心转录因子。Neu5Ac通过抑制IκBα的磷酸化和降解,阻止NF-κB p65亚基向核内转位,从而下调多种促炎基因(如TNF-α、IL-6、iNOS)的转录。这是其抗炎活性的主要分子基础。
* STAT3(信号转导与转录激活因子3):STAT3在调节Th17细胞分化和炎症反应中起关键作用。Neu5Ac被发现能抑制STAT3的磷酸化(Tyr705位点),从而阻断其激活。这有助于抑制Th17介导的自身免疫性炎症。同时,STAT3也参与IL-10介导的抗炎信号,Neu5Ac对STAT3的调控可能影响IL-10的反馈调节环路。
* STAT4:STAT4是IL-12信号通路的关键分子,驱动Th1细胞分化和IFN-γ的产生。Neu5Ac可能通过调节STAT4的磷酸化水平,影响Th1/Th2免疫平衡。
3. 细胞因子与免疫检查点
* IL-2(白细胞介素-2):IL-2是T细胞增殖和存活的关键因子。Neu5Ac被发现能促进T细胞产生IL-2,从而增强适应性免疫应答。
* IL-10(白细胞介素-10):作为主要的抗炎细胞因子,IL-10由Treg细胞等多种细胞分泌。Neu5Ac能显著上调IL-10的表达,这是其诱导免疫耐受和抑制炎症的重要机制。
* TGF-β1(转化生长因子-β1):TGF-β1是Treg细胞分化的关键诱导因子。Neu5Ac通过激活TGF-β1信号,促进初始T细胞向Foxp3⁺ Treg细胞分化,从而建立免疫抑制微环境。
* IFN-γ(干扰素-γ):作为Th1型细胞因子的代表,IFN-γ在抗病毒和抗肿瘤免疫中至关重要。Neu5Ac在特定条件下(如联合抗原刺激)能增强IFN-γ的产生,提升细胞免疫应答。
* CTLA-4(细胞毒性T淋巴细胞相关蛋白4):CTLA-4是T细胞表面的免疫检查点受体,传递抑制性信号。Neu5Ac对CTLA-4表达的影响尚存争议,但部分研究表明其可能通过调节Treg细胞功能间接影响CTLA-4的表达水平。
* FOXP3(叉头框蛋白P3):FOXP3是Treg细胞谱系特异性转录因子。Neu5Ac通过TGF-β1/Smad信号通路诱导FOXP3表达,是驱动Treg细胞分化的核心机制。
综上所述,Neu5Ac并非作用于单一靶点,而是通过“多靶点、多通路”的网络调控模式,精细调节TLR4、STAT3、NF-κB等关键节点,进而影响IL-2、IL-10、TGF-β1、IFN-γ等细胞因子的平衡,最终实现对免疫应答的“双向”或“稳态”调节。
从药物开发的角度审视,Neu5Ac的成药性呈现出“机遇与挑战并存”的特点。
优势方面:
* 高安全性:作为人体内源性物质,其免疫原性低,且Ames试验阴性、无hERG抑制风险,遗传毒性和心脏毒性风险极低。
* 明确的靶点与机制:其与TLR4、Siglecs等免疫受体的相互作用机制研究较为深入,为基于结构的药物设计提供了基础。
* 多效性:兼具免疫调节、抗病毒、神经保护等多种活性,使其具备开发为多功能药物的潜力。
挑战方面:
* 口服生物利用度低:极低的LogP值(-2.58)和高水溶性,导致其难以通过被动扩散穿越肠道上皮细胞膜。口服后,大部分Neu5Ac可能在肠道内被微生物代谢或直接排出体外。因此,口服给药需要高剂量或采用特殊递送系统(如纳米载体、前药设计)来提高吸收。
* 血浆半衰期短:游离的Neu5Ac在血液循环中会被肝脏和肾脏快速清除,半衰期通常较短。这限制了其在体内的有效暴露时间。
* 血脑屏障穿透性差:低BBB渗透性意味着口服或静脉注射的Neu5Ac难以有效进入中枢神经系统,限制了其在神经退行性疾病中的应用。开发能穿越BBB的衍生物或利用鼻内给药途径是潜在解决方案。
* 代谢不稳定性:Neu5Ac在体内可被唾液酸酶(neuraminidase)水解,尤其是在炎症或感染部位,唾液酸酶活性升高,可能导致药物快速失活。
药代动力学特征:
* 吸收:口服吸收差,生物利用度通常低于5%。静脉注射后,分布迅速,主要分布在细胞外液。
* 分布:与血浆蛋白结合率低。由于无法穿越BBB,中枢神经系统中的浓度极低。
* 代谢:主要在肝脏通过去乙酰化、氧化和与葡萄糖醛酸结合等途径代谢。部分Neu5Ac可被重新利用,参与糖蛋白和糖脂的合成。
* 排泄:主要以原形或代谢产物形式通过肾脏排泄。
成药性改良策略:
1. 前药设计:将Neu5Ac的羧基或羟基进行酯化或醚化修饰,提高脂溶性,促进口服吸收。例如,奥司他韦(Tamiflu)就是唾液酸类似物的乙酯前药。
2. 纳米递送系统:将Neu5Ac包裹于脂质体、聚合物纳米粒或介孔二氧化硅纳米粒中,可提高其稳定性、延长循环时间,并实现靶向递送。
3. 结构修饰:开发具有更高受体亲和力或代谢稳定性的Neu5Ac衍生物。例如,在C2位引入疏水基团,或对C9位进行修饰,以增强与病毒NA或Siglec受体的结合能力。
4. 联合用药:与吸收增强剂(如胆酸盐)或P-糖蛋白抑制剂联用,提高口服生物利用度。
基于其独特的药理活性和相对明确的安全性,N-乙酰-神经氨酸在多个临床领域展现出广阔的应用前景。
1. 免疫调节与抗炎治疗
* 自身免疫性疾病:如类风湿关节炎、炎症性肠病、多发性硬化等。通过诱导Treg细胞分化、抑制Th17细胞和NF-κB通路,Neu5Ac或其衍生物有望开发为新型免疫抑制剂,用于控制自身免疫性炎症。
* 过敏性疾病:通过调节Th1/Th2平衡,抑制IgE产生,Neu5Ac可能对过敏性鼻炎、哮喘等疾病具有治疗潜力。
* 移植免疫:其诱导免疫耐受的特性,使其在器官移植后的抗排斥反应治疗中具有应用价值。
2. 抗病毒药物研发
* 广谱抗病毒:鉴于许多病毒利用唾液酸作为受体,Neu5Ac及其类似物可作为广谱病毒进入抑制剂。特别是针对流感病毒、副流感病毒、人偏肺病毒、某些冠状病毒等。
* 抗耐药病毒:开发与现有神经氨酸酶抑制剂(如奥司他韦)作用机制不同的Neu5Ac衍生物,有望克服流感病毒的耐药性问题。
3. 神经退行性疾病
* 阿尔茨海默病:通过抑制Aβ聚集、减轻神经炎症和改善突触可塑性,Neu5Ac或其能穿越BBB的衍生物,可能成为AD的疾病修饰疗法。
* 帕金森病:其抗氧化和抗炎特性,可能对多巴胺能神经元的保护有益。
4. 肿瘤免疫治疗
* 免疫检查点调节:通过阻断肿瘤细胞表面的唾液酸与免疫细胞Siglec受体的相互作用,开发“唾液酸免疫检查点”抑制剂,有望成为新一代抗肿瘤免疫疗法。例如,设计高亲和力的Neu5Ac类似物或抗体,阻断Siglec-7/9介导的免疫抑制信号。
* 联合治疗:将Neu5Ac与PD-1/PD-L1抑制剂或CAR-T细胞疗法联用,可能通过重塑肿瘤微环境,增强抗肿瘤免疫应答。
5. 营养与保健
* 婴幼儿营养:母乳中的Neu5Ac对婴幼儿大脑发育和免疫系统成熟至关重要。在配方奶粉中添加Neu5Ac已成为行业趋势。
* 老年保健:补充Neu5Ac可能有助于维持老年人免疫功能,预防感染和认知衰退。
未来研究方向:
* 结构-活性关系(SAR)研究:系统研究Neu5Ac不同位点修饰对其与TLR4、Siglecs、病毒蛋白等靶点结合活性的影响,指导高效、高选择性衍生物的设计。
* 递送系统优化:开发能高效穿越肠道屏障和血脑屏障的纳米递送系统,解决其药代动力学瓶颈。
* 临床转化研究:开展设计严谨的随机对照临床试验,验证Neu5Ac在特定疾病(如流感、类风湿关节炎、AD)中的疗效和安全性。
* 糖组学与精准医学:结合糖组学技术,分析不同疾病状态下患者体内唾液酸修饰的谱图变化,为Neu5Ac的个性化应用提供依据。
N-乙酰-神经氨酸(燕窝酸)作为自然界中一种古老而精妙的糖分子,其生物学功能远不止于简单的结构组分。从化学结构上看,它是一个高度亲水、带负电的九碳酮糖酸;从药理活性上看,它是一个多功能的免疫调节剂、病毒入侵抑制剂和神经保护因子。其作用机制的核心在于通过与TLR4、STAT3、NF-κB等关键信号节点以及细胞因子网络的精细互动,实现对免疫应答的稳态调控。
尽管其在成药性方面面临口服生物利用度低、代谢不稳定等挑战,但其极高的安全性、明确的靶点和多效性使其成为一个极具吸引力的药物先导化合物。通过前药设计、纳米递送和结构修饰等现代药物化学手段,这些障碍正在被逐步克服。展望未来,随着糖生物学与免疫学的深度融合,以及精准医疗理念的普及,Neu5Ac及其衍生物有望在抗炎、抗病毒、抗肿瘤和神经保护等多个治疗领域取得突破,从一种传统的“滋补佳品”华丽转身为现代医学中的“精准武器”。对它的深入研究,不仅将丰富我们对生命信息传递的理解,更将为人类健康带来新的福祉。

版权所有:© 成都普瑞法科技开发有限公司(2015)备案号:蜀ICP备15035167号-1 客服热线:400-829-7929
技术支持:南京库价















