
产品名称: 磷脂酰乙醇胺(大豆)
英文名: Phosphatidylethanolamine(from Soybean)
中文别名:
英文别名:
Cas 号: 97281-51-1
产品编码:BP2456
分子式: C41H78NO8P
分子量: 744.0336
来源: 大豆
化合物类型:
纯度: 95%~99%
分析方法: HPLC-DAD or/and HPLC-ELSD
鉴定方法: 质谱(Mass), 核磁(NMR)
包装: 棕色小玻璃瓶,按客户需求包装。
存储: 贮存在避光密闭容器中,冷藏或者冷冻长期保存。
样品溶液最好临用新配。如果需要提前配制的话,最好分成独立包装冷冻保存(-20℃以下),临用前再取出解冻,通常可以保存2周。
可以满足克级以上大量需求,详情请咨询。
提供相关图谱和分析方法指导,可以提供包括色谱柱,标准品和分析方法在内的含量测定整体服务包,价格优惠。
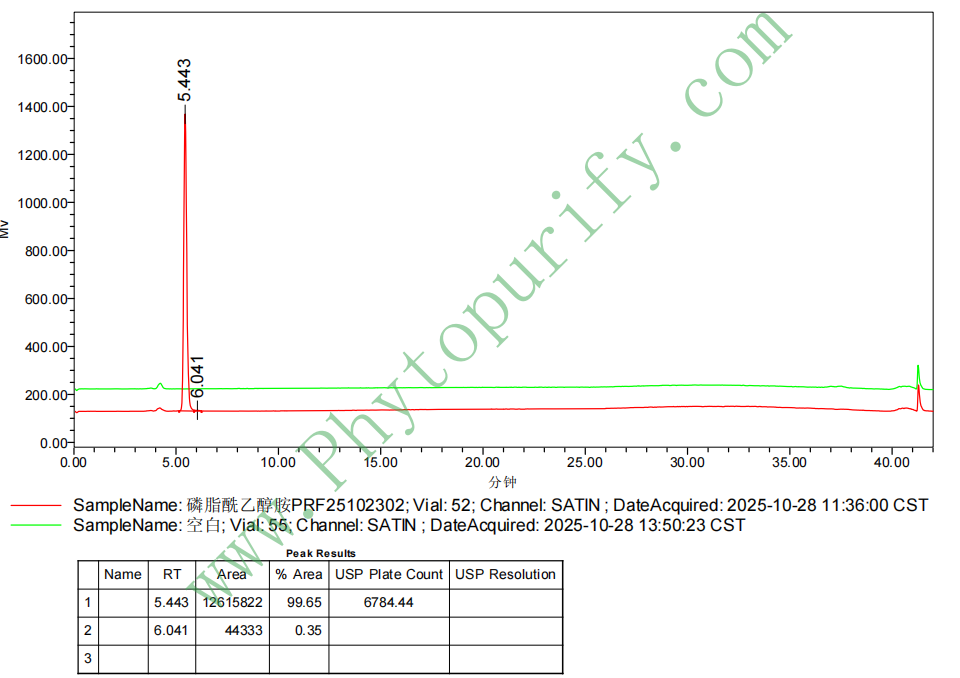
磷脂酰乙醇胺(大豆)的HPLC图谱
储存条件:短期保存 2~8℃,长期保存 -20 ~ -80℃
165.17
否
否
否
否
否
否
否

磷脂是构成生物膜的基本骨架,其结构与功能的多样性决定了细胞生命活动的复杂性。在众多磷脂种类中,磷脂酰乙醇胺(Phosphatidylethanolamine, PE)是仅次于磷脂酰胆碱(Phosphatidylcholine, PC)的第二大丰富的甘油磷脂,广泛存在于哺乳动物、植物及微生物的细胞膜中。PE在维持膜曲率、促进膜融合、参与细胞信号转导以及蛋白质分选等过程中扮演着不可或缺的角色。近年来,随着对细胞自噬(Autophagy)分子机制研究的深入,PE的核心生物学功能被重新定义:它作为自噬相关蛋白Atg8/LC3的脂质锚定物,直接参与自噬体膜的延伸与闭合,成为调控细胞稳态与应激反应的关键脂质分子。
来源于大豆的磷脂酰乙醇胺(Phosphatidylethanolamine (from Soybean),CAS号:97281-51-1)是一种口服活性的天然磷脂。大豆作为全球重要的油料作物,其磷脂提取物是商业化磷脂的主要来源。与动物来源的PE相比,大豆PE具有独特的脂肪酸组成,富含亚油酸(C18:2)和亚麻酸(C18:3)等多不饱和脂肪酸,这赋予了它特殊的物理化学性质和生物活性。研究表明,大豆PE不仅能够增强自噬通量、促进细胞分化、调节脂质滴融合,还在延缓衰老、改善脂质代谢紊乱及维护膜完整性方面展现出潜力。鉴于其良好的安全性(无肝毒性、心脏毒性及遗传毒性)和明确的生物活性,大豆PE正从传统的食品乳化剂和膳食补充剂成分,逐步转变为天然产物药理学和营养药理学领域的研究热点。本文将从化学结构、植物来源、药理活性、作用机制、成药性及临床应用前景等方面,对大豆磷脂酰乙醇胺进行系统的专业综述。
磷脂酰乙醇胺属于甘油磷脂类化合物,其分子结构由三个基本模块构成:一个甘油骨架、两个脂肪酸链(疏水尾)以及一个磷酸乙醇胺极性头基(亲水头)。具体而言,在甘油分子的sn-1和sn-2位点通过酯键连接两条长链脂肪酸,而在sn-3位点则通过磷酸二酯键与乙醇胺基团相连。这种两亲性结构是PE能够形成脂质双分子层、参与膜动态行为的基础。
大豆PE的分子式为C₄₃H₈₂NO₈P,平均分子量约为756.04 Da。其极性表面积(TPSA)为165.17 Ų,较高的TPSA值反映了其极性头基具有较强的氢键形成能力(氢键受体数为9),这与其在水-脂界面上的相互作用特性相符。值得注意的是,大豆PE的脂肪酸组成具有显著特征:与动物来源(如牛脑或蛋黄)的PE富含饱和或单不饱和脂肪酸不同,大豆PE的sn-2位点通常连接着大量的多不饱和脂肪酸,特别是亚油酸(C18:2, ω-6)和α-亚麻酸(C18:3, ω-3)。这种高不饱和度使得大豆PE在常温下具有较低的相变温度,倾向于形成六角形II相(HII相),而非典型的层状相。这一特性赋予了PE独特的“非双层”结构倾向,使其在膜融合、膜曲率调节以及自噬体膜形成等过程中发挥关键作用。
在理化性质方面,大豆PE为淡黄色至棕色的蜡状固体或粉末,具有典型的脂类气味。它不溶于水,但可溶于氯仿、甲醇、乙醇及乙醚等有机溶剂。由于其分子中含有不饱和脂肪酸双键,大豆PE在空气中易发生氧化,需在低温、避光及惰性气体(如氮气)条件下储存。此外,PE的乙醇胺头基在生理pH下带正电荷(pKa约为9.5-10.0),这使其在酸性环境中呈现阳离子特性,而在中性或碱性环境中则表现为两性离子。这种pH依赖的电荷特性影响了PE与蛋白质及其他脂质的静电相互作用。
磷脂酰乙醇胺广泛存在于大豆(Glycine max)的种子中,主要富集于子叶和胚轴组织。在大豆种子中,总磷脂含量约占干重的1.5%-3.0%,其中PE约占磷脂总量的15%-25%,仅次于PC(约30%-40%)。大豆磷脂的组成受品种、种植环境、成熟度及加工工艺的影响而有所波动。
工业上,大豆PE通常作为大豆油精炼过程中的副产品——大豆磷脂(Lecithin)——的组分而被获得。传统的提取工艺主要包括以下几个步骤:首先,通过机械压榨或有机溶剂(正己烷)萃取获得毛油;其次,在毛油脱胶阶段,通过水化作用使磷脂水合沉淀,经离心分离得到湿胶(Wet gum);湿胶经真空干燥后得到粗磷脂。粗磷脂中PE的含量通常为15%-25%,同时含有PC、磷脂酰肌醇(PI)及其他中性脂质。
为了获得高纯度的PE,需要进一步的分离纯化技术。常用的方法包括:
溶剂分馏法:利用PE与PC在低级醇(如乙醇)中的溶解度差异进行分离。PC在乙醇中溶解度较高,而PE溶解度较低,通过控制乙醇浓度和温度,可使PE富集于沉淀相中。该方法操作简便,适合大规模生产,但纯度有限(通常可达60%-70%)。
柱层析法:采用硅胶或氧化铝为固定相,以氯仿-甲醇-水体系为流动相进行梯度洗脱。PE通常在PC之后被洗脱下来。该方法可获得纯度超过95%的PE,但成本较高,主要用于实验室研究或高附加值产品制备。
超临界流体萃取(SFE):使用超临界CO₂作为溶剂,通过添加乙醇等夹带剂选择性萃取PE。该方法绿色环保,避免了有机溶剂残留,但设备投资大,产率受限于PE在超临界CO₂中的溶解度。
酶法改性:利用磷脂酶D(PLD)的转磷脂酰基反应,以PC为底物,在乙醇胺存在下催化合成PE。该方法具有高选择性,可定向生产特定脂肪酸组成的PE。
近年来,大豆PE的药理活性研究取得了显著进展,其生物学效应已超越传统的膜结构组分功能,扩展至细胞信号调控、代谢调节及抗衰老等多个领域。
自噬是一种高度保守的细胞自我降解和回收过程,对于维持细胞内环境稳态、清除受损细胞器及错误折叠蛋白质至关重要。PE是自噬体膜形成的关键脂质。在自噬启动后,Atg8/LC3蛋白被Atg7(E1样酶)和Atg3(E2样酶)催化,通过酰胺键共价连接至PE的乙醇胺头基上,形成LC3-PE复合物(即LC3-II)。LC3-II被锚定在自噬体膜上,驱动膜的延伸、弯曲和闭合。研究表明,外源性补充大豆PE可以增加细胞内PE池的可用性,从而增强LC3脂化效率,提高自噬通量。在神经退行性疾病模型(如亨廷顿舞蹈症、帕金森病)中,大豆PE处理可促进突变蛋白聚集体(如mHTT、α-突触核蛋白)的自噬性清除,减轻细胞毒性。此外,在肝细胞中,PE通过激活自噬途径减少脂质滴积累,改善非酒精性脂肪肝(NAFLD)的病理状态。
PE在细胞分化过程中也发挥重要作用。在神经干细胞和PC12细胞模型中,补充PE可促进神经突生长和神经元标志物(如MAP2、NeuN)的表达。其机制可能与PE调节膜流动性、影响生长因子受体(如TrkA)在膜上的定位和信号转导有关。在成骨细胞分化中,PE通过激活Wnt/β-catenin信号通路,上调Runx2和Osterix等成骨关键转录因子的表达,促进矿化结节形成。这些发现提示PE在组织再生和修复中具有潜在应用价值。
PE是线粒体膜和内质网膜的重要组成部分。在线粒体中,PE参与呼吸链复合物的组装和功能维持。大豆PE的补充可改善线粒体膜电位,减少活性氧(ROS)产生,保护线粒体功能。在肝脏中,PE通过影响极低密度脂蛋白(VLDL)的组装和分泌,调节全身脂质代谢。PE缺乏会导致内质网应激和脂质代谢紊乱,而外源性PE补充可逆转这些效应。此外,PE通过其非双层结构倾向,在膜融合事件(如囊泡运输、病毒入侵)中发挥关键作用,维护膜的动态平衡。
衰老过程中,细胞膜中PE含量下降,伴随膜流动性降低和氧化应激增加。大豆PE富含多不饱和脂肪酸,补充后可恢复膜脂肪酸组成,改善膜功能。在秀丽隐杆线虫(C. elegans)衰老模型中,PE处理显著延长了平均寿命,并提高了应激抵抗能力。在哺乳动物细胞中,PE通过抑制NF-κB通路,减少促炎细胞因子(如TNF-α、IL-6)的释放,表现出抗炎活性。这些效应部分归因于PE对自噬的激活,因为自噬本身具有抑制炎症小体活化的功能。
大豆PE的药理活性根植于其独特的分子相互作用网络,主要涉及以下几个关键机制和靶点:
这是PE最核心的分子功能。在自噬过程中,PE作为底物参与Atg8/LC3的脂化修饰。具体而言,Atg4蛋白酶首先切割pro-LC3的C端,暴露甘氨酸残基(LC3-I)。随后,LC3-I在Atg7(E1)和Atg3(E2)的催化下,其C端甘氨酸与PE的乙醇胺头基形成酰胺键,生成LC3-II。LC3-II被招募至自噬体膜,作为膜延伸的支架蛋白。大豆PE的补充增加了局部PE浓度,提高了LC3脂化效率,从而增强自噬通量。这一过程受Atg3与PE的亲和力调控,而PE的脂肪酸组成(尤其是链长和不饱和度)会影响其与Atg3的结合效率。
PE因其较小的极性头基和较大的疏水体积,倾向于形成负曲率膜结构(如六角形II相)。这种特性使PE成为膜融合和膜弯曲的关键调节因子。在自噬体形成过程中,PE富集于自噬体膜的弯曲部位,促进膜的延伸和闭合。此外,PE通过调节SNARE蛋白复合物的组装,参与囊泡与靶膜的融合过程。大豆PE的高不饱和度增强了其形成非双层结构的能力,从而更有效地促进膜融合事件。
PE是线粒体膜中含量最丰富的磷脂之一(约占线粒体磷脂的20%-30%)。在线粒体内膜中,PE与心磷脂(Cardiolipin)协同作用,维持呼吸链复合物(尤其是复合物I、III和IV)的稳定性和活性。PE通过与线粒体蛋白(如ADP/ATP转位酶ANT)的直接相互作用,调节线粒体通透性转换孔(mPTP)的开放。大豆PE的补充可改善线粒体呼吸效率,减少电子泄漏和ROS产生。
PE及其代谢产物(如溶血磷脂酰乙醇胺LPE)可作为信号分子,激活特定的受体或信号通路。例如,PE通过调节膜微域(脂筏)的组成,影响受体酪氨酸激酶(如EGFR、TrkA)的聚集和激活。此外,PE代谢产生的乙醇胺可进入CDP-乙醇胺途径,影响磷脂合成和细胞增殖。在转录水平上,PE通过调节内质网应激传感器(如IRE1、PERK)的活性,影响未折叠蛋白反应(UPR)和脂质代谢相关基因的表达。
从药物开发的角度看,大豆PE具有一系列理想的成药性特征,但也面临若干挑战。
根据现有毒理学数据,大豆PE表现出良好的安全性。Ames试验结果为阴性,表明其无遗传毒性。在hERG抑制试验中未观察到心脏毒性风险,且无明确的肝毒性信号。这些数据支持其作为口服制剂的安全性。然而,需要注意的是,PE在体内代谢后产生的溶血磷脂和游离脂肪酸,在高剂量下可能引起细胞膜扰动,需进行长期毒性研究以确定安全剂量范围。
大豆PE的口服生物利用度受其脂溶性本质和胃肠道环境的影响。口服后,PE在肠道中首先被磷脂酶A2(PLA2)水解,释放sn-2位脂肪酸,生成溶血磷脂酰乙醇胺(LPE)。LPE可通过被动扩散或转运蛋白(如MFSD2A)被肠上皮细胞吸收。在细胞内,LPE被重新酰化生成PE,或以LPE形式进入淋巴循环。由于PE的分子量较大(>700 Da)且TPSA较高(>160 Ų),其血脑屏障穿透能力极低(BBB: No),这限制了其在中枢神经系统疾病中的直接应用。PE主要经肝脏代谢,通过磷脂酶和酯酶水解,最终产物进入β-氧化或磷脂再循环途径。
为改善PE的口服生物利用度和靶向性,多种制剂策略正在探索中。脂质体技术可将PE包裹于磷脂双分子层中,提高其水分散性和稳定性。纳米乳剂和自微乳化给药系统(SMEDDS)可促进PE在胃肠道的溶解和吸收。此外,通过将PE与聚乙二醇(PEG)或其他靶向配体偶联,可实现PE的靶向递送,例如靶向肝脏或肿瘤组织。
基于大豆PE的多效药理活性,其在以下疾病领域的应用前景值得关注:
非酒精性脂肪肝病(NAFLD)和非酒精性脂肪性肝炎(NASH)是日益严峻的全球健康问题。PE通过增强自噬通量、促进脂质滴降解、改善线粒体功能,有望成为NAFLD/NASH的辅助治疗手段。临床前研究已证实,PE补充可减轻肝脏脂肪变性、炎症和纤维化。未来需要开展随机对照临床试验,验证其在NAFLD患者中的疗效和安全性。
尽管PE难以穿透血脑屏障,但通过鼻内给药或脂质体包裹等途径,仍有可能实现中枢递送。在阿尔茨海默病(AD)模型中,PE通过促进Aβ的自噬性清除和抑制tau蛋白磷酸化,改善认知功能。在帕金森病(PD)中,PE保护多巴胺能神经元免受MPTP诱导的毒性。鉴于自噬功能障碍是神经退行性疾病的共同病理特征,PE作为自噬增强剂具有广阔的应用前景。
PE在延缓衰老方面的潜力已引起化妆品和营养保健品行业的关注。局部应用PE可改善皮肤屏障功能,减少皱纹形成。口服PE补充剂可能通过系统性抗炎和抗氧化作用,延缓器官衰老。然而,目前缺乏大规模的人体研究数据,其抗衰老效果仍需进一步验证。
自噬在肿瘤发生发展中具有双重作用:在肿瘤早期,自噬抑制肿瘤生长;在晚期,自噬促进肿瘤细胞存活。因此,PE作为自噬增强剂的应用需谨慎。在某些特定肿瘤类型(如肝癌、胰腺癌)中,PE可能通过诱导自噬性细胞死亡或增强化疗敏感性而发挥抗肿瘤作用。此外,PE作为膜组分,可影响肿瘤细胞对化疗药物的摄取和耐药性。
磷脂酰乙醇胺(大豆)作为一种天然、安全且具有多重生物活性的磷脂分子,正从传统的膜结构组分转变为具有重要药理价值的天然产物。其通过参与Atg8/LC3脂化、调节膜曲率、维护线粒体功能等机制,在自噬调控、细胞分化、脂质代谢及抗衰老等方面展现出独特的生物学效应。大豆来源的PE因其丰富的多不饱和脂肪酸组成,在生物活性上可能优于动物来源的PE。尽管其口服生物利用度和血脑屏障穿透性存在局限,但通过先进的制剂技术有望克服这些障碍。未来,随着对PE分子机制理解的深入以及临床转化研究的推进,大豆PE有望在代谢性疾病、神经退行性疾病及抗衰老等领域发挥更大的治疗潜力。然而,从基础研究到临床应用的道路仍充满挑战,包括明确最佳剂量、建立标准化提取工艺、开展长期安全性评价以及设计严谨的临床试验。磷脂酰乙醇胺(大豆)的研究,不仅深化了我们对脂质生物学功能的认识,也为开发新型天然药物和功能性食品提供了重要方向。

版权所有:© 成都普瑞法科技开发有限公司(2015)备案号:蜀ICP备15035167号-1 客服热线:400-829-7929
技术支持:南京库价















