
产品名称: 二氢大豆苷元
英文名: DihydroDaidzein
中文别名: (S)-二氢大豆苷元
英文别名: 17238-05-0, (±)-Dihydrodaidzein;S-2,3-dihydro-7-hydroxy-3-(4-hydroxyphenyl)-4H-1-Benzopyran-4-one
Cas 号: 879559-75-8
产品编码:BP3946
分子式: C15H12O4
分子量: 256.257
来源:
物理性状:
化合物类型:
纯度: 95%~99%
分析方法: HPLC-DAD or/and HPLC-ELSD
鉴定方法: 质谱(Mass), 核磁(NMR)
包装: 棕色小玻璃瓶,按客户需求包装。
存储: 贮存在避光密闭容器中,冷藏或者冷冻长期保存。
样品溶液最好临用新配。如果需要提前配制的话,最好分成独立包装冷冻保存(-20℃以下),临用前再取出解冻,通常可以保存2周。
可以满足克级以上大量需求,详情请咨询。
提供相关图谱和分析方法指导,可以提供包括色谱柱,标准品和分析方法在内的含量测定整体服务包,价格优惠。
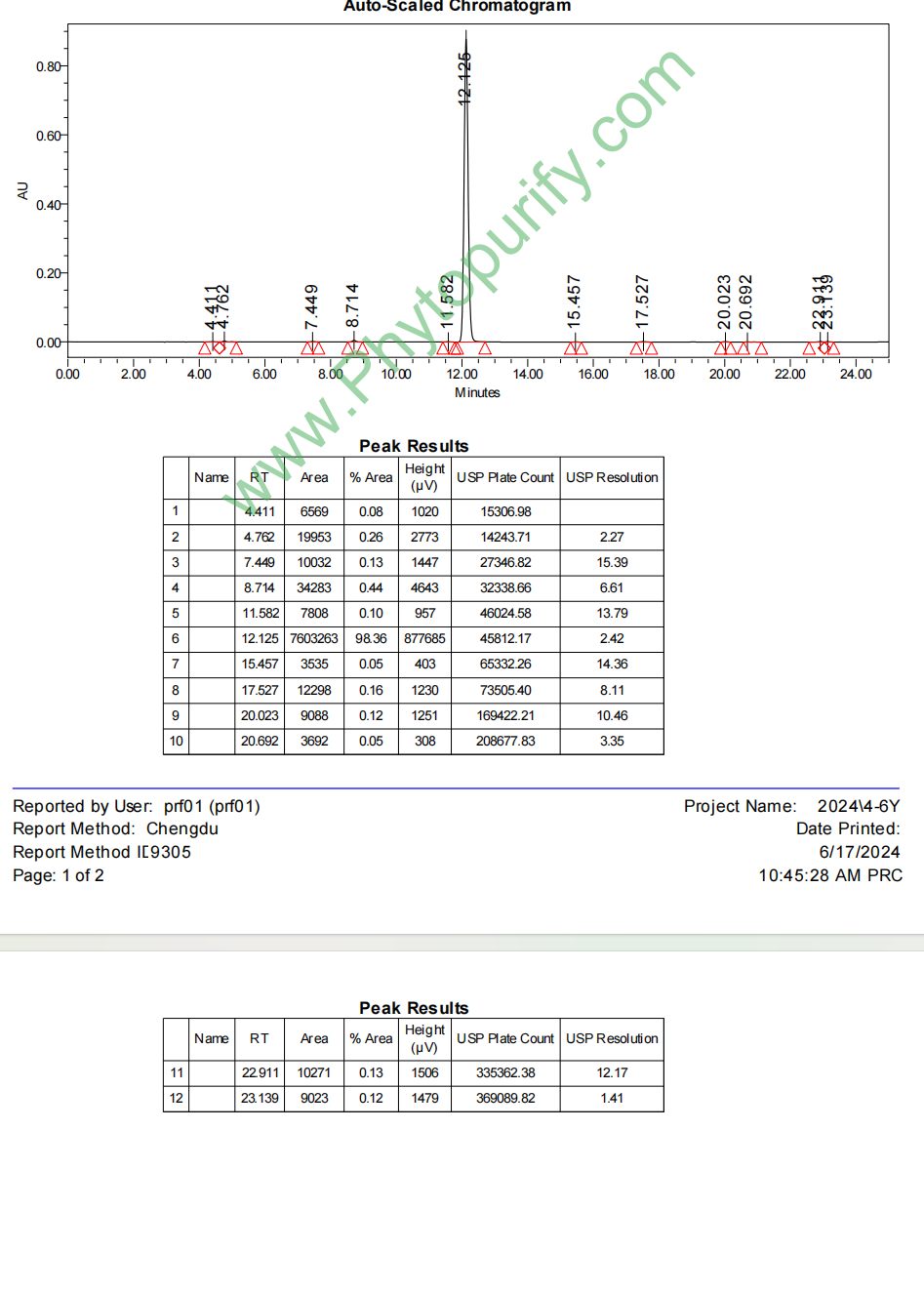
二氢大豆苷元的HPLC图谱
储存条件:短期保存 2~8℃,长期保存 -20 ~ -80℃
66.76
2.38
2.34
0.27
3.19
14.97
低
88.25
2.72
否
否
否
否
有
无
0.0
是
是
是
是

天然产物作为药物发现的重要源泉,在人类健康维护和疾病治疗中扮演着不可或缺的角色。在众多具有生物活性的天然产物中,植物雌激素(Phytoestrogens)因其与内源性雌激素17β-雌二醇的结构相似性,能够与雌激素受体(Estrogen Receptors, ERs)相互作用,从而在激素相关疾病、代谢性疾病及癌症的预防与治疗中展现出巨大潜力。大豆异黄酮(Soy Isoflavones)是研究最为深入的一类植物雌激素,主要包括大豆苷元(Daidzein)、染料木素(Genistein)和黄豆黄素(Glycitein)。然而,这些异黄酮在体内经肠道微生物代谢后,会转化为一系列具有独特生物活性的代谢产物,其中,二氢大豆苷元(Dihydrodaidzein, DHD)便是大豆苷元代谢途径中的关键中间体。
二氢大豆苷元,英文名Dihydrodaidzein,CAS号为879559-75-8,化学名为4',7-二羟基异黄烷酮(4',7-Dihydroxyisoflavanone)。它并非植物直接大量合成的原始成分,而是大豆苷元在哺乳动物肠道内,经由特定菌群介导的还原反应生成的代谢产物。这一转化过程是理解大豆异黄酮个体差异生物效应的关键。与母体化合物大豆苷元相比,二氢大豆苷元在结构上发生了显著变化,其异黄酮骨架中的C2-C3双键被还原,形成了一个手性中心,从而可能赋予其不同的立体化学特性和生物活性。作为“最著名的可食用性植物雌激素之一”,二氢大豆苷元不仅继承了与雌激素受体结合的能力,更因其独特的代谢中间体身份,在调节雌激素信号通路、抗氧化、抗炎以及抗肿瘤等方面展现出独特的研究价值。
近年来,随着对肠道微生物组(Gut Microbiome)与宿主健康相互作用认识的不断深入,二氢大豆苷元作为连接膳食异黄酮与生物活性代谢物(如雌马酚,Equol)的桥梁分子,其重要性日益凸显。它不仅自身具有药理活性,更是合成具有更强雌激素活性的终末代谢产物雌马酚的前体。因此,深入解析二氢大豆苷元的化学性质、生物来源、药理作用及其分子机制,对于阐明大豆食品的健康功效、开发基于肠道微生物调控的精准营养策略以及设计新型植物雌激素类药物具有重要的科学意义和临床应用价值。本文旨在对二氢大豆苷元的研究现状进行系统综述,涵盖其化学结构、理化性质、来源、药理活性、作用机制、成药性及临床应用前景,以期为该领域的深入研究提供参考。
二氢大豆苷元的化学结构是其生物学功能的基础。从化学分类上看,它属于异黄烷酮(Isoflavanone),是大豆苷元(一种异黄酮,Isoflavone)的C2-C3双键被还原后的产物。其系统命名为4',7-二羟基异黄烷酮。该分子由一个二苯基丙烷骨架构成,其中B环连接在C3位,这与异黄酮(B环连接在C3位)和异黄烷(Isoflavan,B环连接在C3位且无双键和羰基)的结构特征相呼应。二氢大豆苷元的核心结构是一个二氢色原酮(Chromanone)环系,C4位为羰基,C2和C3位为单键,这使得C2和C3成为两个手性中心。因此,二氢大豆苷元存在四种可能的立体异构体:(2R,3R)-、(2R,3S)-、(2S,3R)-和(2S,3S)-二氢大豆苷元。天然来源或生物转化产生的二氢大豆苷元通常以特定的对映体或非对映体混合物形式存在,其立体构型对其与生物大分子(如雌激素受体)的结合亲和力及后续生物活性具有决定性影响。例如,研究表明,不同构型的二氢大豆苷元在雌激素活性上可能存在显著差异。
在理化性质方面,二氢大豆苷元(分子式C₁₅H₁₂O₄)的分子量为256.2570 g/mol。其脂水分配系数LogP为2.3770,表明该化合物具有适中的亲脂性,这使其能够较容易地穿过细胞膜,但也限制了其在水相环境中的溶解度。其水溶性(Water Solubility)参数为0.2725 mg/mL,属于微溶范畴,这与其分子结构中存在两个酚羟基(4'-OH和7-OH)以及一个羰基(C=O)有关,这些极性基团能够与水分子形成氢键,但整体非极性的芳香环骨架仍占主导。拓扑极性表面积(Topological Polar Surface Area, TPSA)为66.7600 Ų。TPSA是预测药物口服吸收和血脑屏障穿透能力的重要参数。通常,TPSA小于60 Ų的分子易于穿透血脑屏障,而大于140 Ų的分子则难以穿透。二氢大豆苷元的TPSA为66.76 Ų,略高于60 Ų的阈值,这与其被预测为“血脑屏障穿透性低”的特性相符。这意味着二氢大豆苷元在中枢神经系统中的分布可能有限,其药理作用主要集中在外周组织。此外,该分子含有两个酚羟基,使其具有一定的酸性(pKa约在7-10之间),在生理pH条件下部分电离,这也会影响其溶解度和与蛋白质的结合。其紫外吸收特征主要来源于A环和B环的芳香系统,在约280 nm和310 nm处有特征吸收峰,可用于其定性和定量分析。
与大豆苷元和染料木素等直接由植物合成的大豆异黄酮不同,二氢大豆苷元在植物中的含量极低,通常不被视为植物直接产生的次生代谢产物。其主要来源是哺乳动物(包括人类)体内的肠道微生物代谢。因此,二氢大豆苷元的“来源”更准确地说是“生物转化来源”。
肠道微生物转化:当人们摄入富含大豆苷元的食物(如豆腐、豆浆、豆粉等)后,大豆苷元在小肠被吸收,但大部分以糖苷形式(如大豆苷,Daidzin)进入结肠。在结肠中,肠道菌群分泌的β-葡萄糖苷酶(β-Glucosidase)首先将大豆苷水解为苷元——大豆苷元。随后,特定的肠道细菌,如某些乳酸菌(Lactobacillus spp.)、双歧杆菌(Bifidobacterium spp.)以及梭菌(Clostridium spp.)等,通过其表达的还原酶(如异黄酮还原酶,Isoflavone Reductase),将大豆苷元C2-C3双键还原,生成二氢大豆苷元。这一还原反应是立体选择性的,不同菌株可能产生不同构型的二氢大豆苷元。二氢大豆苷元是进一步代谢为雌马酚(Equol)或去氧甲基安哥拉紫檀素(O-Desmethylangolensin, O-DMA)的关键前体。因此,个体肠道菌群的组成差异直接决定了二氢大豆苷元的生成效率及其后续代谢途径,这也是为什么人群中存在“雌马酚生产者”和“非生产者”表型的重要原因。
提取与分离方法:由于二氢大豆苷元在自然界中并非大量存在,其获取主要依赖于化学合成或生物转化法。
化学合成法:这是获得标准品和进行大规模研究的主要方法。通常以大豆苷元为起始原料,通过催化氢化(如使用钯碳、铂等催化剂在氢气条件下)选择性还原C2-C3双键来合成二氢大豆苷元。该方法的挑战在于控制反应的立体选择性,以获得单一构型的产物。通过使用手性催化剂或手性辅助试剂,可以实现对特定对映体的不对称合成。合成的产物需要通过柱色谱、重结晶或高效液相色谱(HPLC)进行纯化。
生物转化法:利用特定的微生物(如工程化的Escherichia coli或天然产二氢大豆苷元的肠道菌株)在发酵罐中,以大豆苷元为底物进行生物转化。这种方法条件温和,环境友好,且可能获得与体内代谢一致的天然构型产物。发酵液经过离心、过滤后,通过液-液萃取(如使用乙酸乙酯)或固相萃取(SPE)富集目标产物,再经制备型HPLC进行分离纯化。
从生物样品中提取:为了研究体内代谢,可以从服用大豆制品后的动物或人类血液、尿液或粪便中提取二氢大豆苷元。样品通常需要经过酶解(如使用β-葡萄糖醛酸酶/硫酸酯酶)以水解其与葡萄糖醛酸或硫酸的结合物,然后通过液-液萃取或SPE进行富集,最后用HPLC或液相色谱-质谱联用(LC-MS/MS)进行定性和定量分析。
分析检测方法:二氢大豆苷元的分析检测主要依赖于高效液相色谱(HPLC)结合紫外检测器(UV)或质谱检测器(MS)。LC-MS/MS因其高灵敏度和高特异性,成为生物样品中痕量二氢大豆苷元定量的金标准。手性色谱柱(如Chiralcel OD-H等)可用于分离和鉴定二氢大豆苷元的立体异构体。
二氢大豆苷元作为大豆苷元的关键代谢产物,其药理活性研究虽不如母体化合物广泛,但已揭示出其在多个生理病理过程中的潜在干预作用。
雌激素样活性:这是二氢大豆苷元最核心的药理活性。作为植物雌激素,它能与雌激素受体ERα和ERβ结合。研究表明,二氢大豆苷元对ERβ具有相对更高的亲和力,这与许多其他植物雌激素(如染料木素)的特性相似。其雌激素活性通常弱于内源性雌二醇,但强于其前体大豆苷元。这种选择性ERβ激动作用被认为与许多健康益处相关,例如在心血管系统中,ERβ激活可介导血管舒张、抗炎和抗动脉粥样硬化效应;在骨骼系统中,ERβ激动剂可促进成骨细胞分化,抑制破骨细胞活性,从而预防骨质疏松;在神经系统,ERβ的激活具有神经保护作用。二氢大豆苷元能够刺激MCF-7乳腺癌细胞(ER阳性)的增殖,表现出弱雌激素效应,但其效应远低于雌二醇,且在高浓度下可能表现出抗雌激素活性。
抗氧化活性:二氢大豆苷元分子结构中的两个酚羟基(4'-OH和7-OH)是其发挥抗氧化活性的关键基团。它们可以作为氢原子供体,清除自由基(如DPPH自由基、ABTS自由基、羟基自由基等),从而阻断脂质过氧化链式反应。研究表明,二氢大豆苷元在体外实验中表现出显著的抗氧化能力,能够保护细胞免受氧化应激诱导的损伤。其抗氧化活性可能与其雌激素活性协同,共同发挥对心血管和神经系统的保护作用。
抗炎活性:慢性炎症是多种疾病(如心血管疾病、糖尿病、癌症)的共同病理基础。二氢大豆苷元被证实具有抗炎作用。在脂多糖(LPS)刺激的巨噬细胞模型中,二氢大豆苷元能够显著抑制促炎细胞因子(如TNF-α、IL-6、IL-1β)和一氧化氮(NO)的产生。其机制可能与抑制核因子κB(NF-κB)信号通路的激活有关。NF-κB是炎症反应的核心转录因子,抑制其活性可下调多种炎症介质的表达。
抗肿瘤活性:二氢大豆苷元的抗肿瘤作用备受关注,尤其是在激素依赖性癌症(如乳腺癌、前列腺癌)方面。其作用机制复杂,可能涉及多种途径:
代谢调节作用:作为雌马酚的前体,二氢大豆苷元在调节糖脂代谢方面也显示出潜力。研究表明,二氢大豆苷元及其代谢产物雌马酚能够改善胰岛素敏感性,促进葡萄糖摄取,并调节脂肪细胞的分化和脂质代谢。这些作用可能通过激活PPARγ(过氧化物酶体增殖物激活受体γ)等核受体来实现,从而对2型糖尿病和肥胖症产生有益影响。
二氢大豆苷元的药理活性源于其与多种分子靶点的相互作用,其机制网络复杂,涵盖了从细胞膜受体到核内转录因子的多个层面。
雌激素受体(ER)介导的信号通路:
NF-κB信号通路:二氢大豆苷元的抗炎作用主要归因于其对NF-κB通路的抑制。在静息状态下,NF-κB与抑制蛋白IκB结合,以无活性形式存在于细胞质中。当受到LPS、TNF-α等炎症刺激时,IκB激酶(IKK)被激活,磷酸化IκB,导致其泛素化降解。释放的NF-κB转位入核,启动促炎基因(如TNF-α, IL-6, COX-2, iNOS)的转录。二氢大豆苷元可通过抑制IKK的活性,或直接干扰NF-κB与DNA的结合,从而阻断这一过程,发挥抗炎效应。
AMPK/mTOR信号通路:AMPK是细胞能量代谢的关键传感器。二氢大豆苷元被报道可激活AMPK,可能通过增加AMP/ATP比率或直接作用于AMPK的调控亚基。活化的AMPK磷酸化并抑制下游的mTORC1复合物。mTORC1是细胞生长和增殖的核心调控者。抑制mTORC1活性可导致蛋白质合成减少、细胞周期阻滞和自噬诱导。这一机制被认为是二氢大豆苷元发挥抗肿瘤和代谢调节作用的重要途径。
抗氧化防御系统:二氢大豆苷元不仅直接清除自由基,还能通过激活核因子E2相关因子2(Nrf2)信号通路,增强细胞内源性抗氧化防御能力。Nrf2是一种转录因子,在氧化应激下从Keap1蛋白释放,转位入核,与抗氧化反应元件(ARE)结合,启动一系列抗氧化酶(如HO-1, NQO1, SOD, CAT)和II相解毒酶的表达。二氢大豆苷元可能通过修饰Keap1上的半胱氨酸残基,或激活上游激酶(如PI3K, MAPK),来促进Nrf2的核转位。
其他潜在靶点:
将二氢大豆苷元开发为候选药物,需要对其成药性进行系统评估。根据提供的参数,并结合其药代动力学特性,可以对其药物开发潜力进行分析。
成药性参数分析:
- 分子量 (256.2570):符合“Lipinski五规则”中分子量小于500的要求,表明其具有良好的口服吸收潜力。
- LogP (2.3770):脂溶性适中,介于1-3之间,是理想的药物候选范围。适中的LogP有利于药物在体内的跨膜转运和分布。
- TPSA (66.7600 Ų):小于140 Ų,预示其口服吸收良好。同时,该值略高于60 Ų,表明其血脑屏障穿透性低,这既是优点也是缺点。优点在于可减少中枢神经系统副作用,缺点在于限制了其在脑部疾病(如阿尔茨海默病)中的应用。
- 水溶性 (0.2725 mg/mL):属于微溶范畴。虽然不理想,但通过制剂技术(如固体分散体、环糊精包合物、纳米乳剂等)可以显著提高其生物利用度。
- hERG抑制 (否):这是一个非常积极的信号。hERG(人ether-à-go-go相关基因)钾通道抑制是导致药物性心脏QT间期延长和致命性心律失常(如尖端扭转型室性心动过速)的主要原因。二氢大豆苷元无hERG抑制活性,大大降低了其心脏毒性风险。
- Ames试验 (0.0):Ames试验用于检测化合物的致突变性。结果为0.0,表明其在测试条件下无致突变性,遗传毒性风险低,这是药物开发早期安全性评价的重要优势。
药代动力学特性:
- 吸收:二氢大豆苷元主要在结肠由肠道菌群生成,其本身并非直接口服吸收的主要形式。口服大豆苷元后,其吸收和代谢存在显著的个体差异,主要取决于肠道菌群的组成。生成的二氢大豆苷元一部分被结肠上皮细胞吸收,进入门静脉循环。其吸收速率和程度受肠道菌群活性、肠道pH值、食物基质等因素影响。
- 分布:吸收后,二氢大豆苷元主要与血浆蛋白(尤其是白蛋白)结合。其适中的LogP和TPSA表明其组织分布广泛,主要分布在外周组织,如肝脏、肾脏、乳腺、前列腺、骨骼等。由于血脑屏障穿透性低,其在中枢神经系统的浓度较低。
- 代谢:二氢大豆苷元在体内经历广泛的II相代谢,主要是在肝脏和肠道黏膜中与葡萄糖醛酸或硫酸结合,形成水溶性更强的结合物,便于从尿液和胆汁中排泄。这些结合物可经肠肝循环重新进入肠道,被肠道菌群的β-葡萄糖醛酸酶水解,重新释放出游离的二氢大豆苷元,从而延长其在体内的滞留时间。更重要的是,二氢大豆苷元是雌马酚的直接前体,在特定肠道菌群的作用下,其C4位羰基可被进一步还原为羟基,生成雌马酚。
- 排泄:二氢大豆苷元及其代谢产物(包括雌马酚、O-DMA以及各种结合物)主要通过尿液和粪便排泄。其半衰期和清除率因个体肠道菌群差异而不同。
成药性评价总结:二氢大豆苷元具备良好的成药性基础。其分子量、脂溶性、无hERG抑制和Ames试验阴性等特性均为其药物开发提供了有利条件。主要挑战在于其水溶性较差和口服生物利用度高度依赖于个体肠道菌群。未来的药物开发策略可以包括:1)开发前药,以提高其口服吸收和生物利用度;2)设计成缓释制剂,以模拟其在结肠的持续生成;3)与益生菌或益生元联用,以调控肠道菌群,优化其体内转化;4)开发其特定立体异构体,以增强靶点选择性和药效。
基于二氢大豆苷元独特的药理活性和良好的成药性,其在多个疾病领域的应用前景广阔,但也面临着诸多挑战。
临床应用前景:
1. 更年期综合征的替代疗法:二氢大豆苷元作为植物雌激素,其选择性ERβ激动作用使其成为缓解更年期症状(如潮热、盗汗、情绪波动)的潜在替代疗法。与传统的激素替代疗法(HRT)相比,其副作用风险(如乳腺癌、血栓风险)可能更低。开发基于二氢大豆苷元或其前体(如大豆苷元)与特定益生菌组合的膳食补充剂,有望为更年期女性提供更安全有效的选择。
骨质疏松症的预防与治疗:通过激活成骨细胞中的ERβ,二氢大豆苷元可促进骨形成,同时抑制破骨细胞活性,减少骨吸收。其抗炎和抗氧化活性也有助于维持骨微环境的稳态。临床前研究已显示出其抗骨质疏松的潜力,未来可开发为治疗或预防绝经后骨质疏松症的药物或功能食品。
心血管疾病的辅助治疗:二氢大豆苷元通过改善血管内皮功能(激活eNOS,促进NO生成)、抗炎(抑制NF-κB)、抗氧化(激活Nrf2)和调节脂质代谢(激活PPARγ)等多重机制,发挥心血管保护作用。它可作为他汀类等降脂药物的辅助治疗,或用于心血管疾病的一级预防。
代谢综合征的干预:鉴于其在改善胰岛素敏感性、调节糖脂代谢方面的作用,二氢大豆苷元有望用于2型糖尿病和肥胖症的干预。其AMPK激活作用是其发挥代谢调节效应的核心机制之一。
激素依赖性癌症的化学预防:对于乳腺癌和前列腺癌高风险人群,长期摄入富含二氢大豆苷元或其前体的食物,可能通过其弱雌激素/抗雌激素活性以及非激素依赖性机制(如诱导凋亡、抑制血管生成),起到化学预防作用。然而,对于已确诊的ER阳性乳腺癌患者,其使用需极为谨慎,因为其弱雌激素活性理论上可能刺激肿瘤生长。
面临的挑战与未来展望:
1. 个体差异问题:最大的挑战在于二氢大豆苷元的体内生成高度依赖于个体肠道菌群。人群中“雌马酚生产者”的比例在不同种族和地区差异很大(约30-50%)。如何确保非生产者也能从二氢大豆苷元治疗中获益,是亟待解决的问题。未来的方向是开发“精准益生菌”或“合成生物学菌株”,使其能够在所有个体肠道中稳定地将大豆苷元转化为二氢大豆苷元。
立体异构体的研究:目前对二氢大豆苷元的研究多使用外消旋混合物。不同立体异构体的药理活性、代谢途径和安全性可能存在显著差异。未来需要系统研究单一构型二氢大豆苷元的生物活性,并开发其高纯度、高立体选择性的合成方法。
临床转化研究:目前的研究多停留在体外和动物实验层面。亟需设计严谨、大规模、随机对照的临床试验,以验证二氢大豆苷元在人体中的有效性、安全性和最佳剂量。需要开发可靠的生物标志物(如血液或尿液中二氢大豆苷元及其代谢物的浓度)来评估其暴露水平和药效。
制剂与递送系统:为解决其水溶性差和口服生物利用度低的问题,需要开发先进的药物递送系统,如脂质体、纳米粒、磷脂复合物等,以提高其口服吸收和靶向递送能力。
安全性评估:尽管Ames试验阴性,但长期使用的安全性,特别是对激素敏感组织(如乳腺、子宫)的潜在影响,仍需通过长期毒理学研究和上市后监测来全面评估。
二氢大豆苷元,作为大豆苷元在肠道微生物作用下的关键代谢产物,已超越其作为简单中间体的角色,展现出作为多靶点、多功能天然活性分子的巨大潜力。其独特的化学结构赋予了它适中的亲脂性和良好的安全性特征(无hERG抑制、无致突变性),使其成为药物开发的理想候选骨架。从雌激素受体信号通路到AMPK、NF-κB、Nrf2等关键调控网络,二氢大豆苷元通过复杂的分子机制,在更年期综合征、骨质疏松、心血管疾病、代谢综合征乃至癌症的预防与治疗中展现出令人瞩目的应用前景。
然而,从实验室研究到临床应用的道路依然充满挑战。个体肠道菌群的巨大差异是其药代动力学和药效学不确定性的根源,也是实现精准治疗的主要障碍。未来的研究重点应聚焦于:深入解析其不同立体异构体的构效关系;开发能够稳定、高效地将大豆苷元转化为二氢大豆苷元的工程菌或益生菌组合;利用先进的制剂技术克服其溶解度和生物利用度瓶颈;并通过高质量的临床研究验证其确切疗效和长期安全性。
总而言之,二氢大豆苷元是连接膳食、肠道微生物与宿主健康的一座重要桥梁。对它的深入研究,不仅将深化我们对大豆食品健康功效分子机制的理解,更有望催生出一类基于肠道微生物调控的新型植物雌激素药物或功能食品,为人类健康事业做出贡献。随着系统生物学、合成生物学和精准医学的不断发展,二氢大豆苷元的临床转化前景值得期待。

版权所有:© 成都普瑞法科技开发有限公司(2015)备案号:蜀ICP备15035167号-1 客服热线:400-829-7929
技术支持:南京库价















