
产品名称: 肿柄菊内酯A
英文名: Tagitinin A
中文别名:
英文别名:
Cas 号: 59979-61-2
产品编码:SBP00186
分子式: C19H28O7
分子量: 368.426
来源: 肿柄菊
纯度: 95%~99%
分析方法: HPLC-DAD or/and HPLC-ELSD
鉴定方法: 质谱(Mass), 核磁(NMR)
包装: 棕色小玻璃瓶,按客户需求包装。
存储: 贮存在避光密闭容器中,冷藏或者冷冻长期保存。
样品溶液最好临用新配。如果需要提前配制的话,最好分成独立包装冷冻保存(-20℃以下),临用前再取出解冻,通常可以保存2周。
可以满足克级至公斤级大量需求,详情请咨询。
提供相关图谱和分析方法指导,可以提供包括色谱柱,对照品和分析方法在内的含量测定整体服务包,价格优惠。
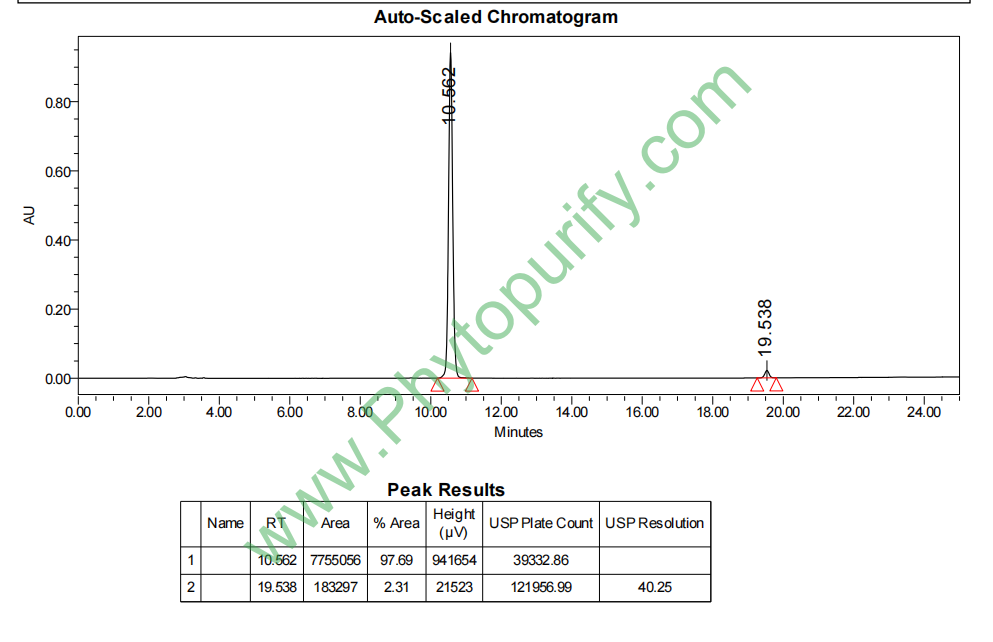
肿柄菊内酯A的HPLC图谱
储存条件:短期保存 2~8℃,长期保存 -20 ~ -80℃
102.29
0.80
0.80
0.87
1.86
1.70
高
53.37
6.07
否
否
否
否
有
无
0.0
是
否
是
是

天然产物一直是药物发现与开发的重要源泉,尤其在抗肿瘤和代谢性疾病治疗领域,其化学多样性和独特的生物活性为现代药物化学提供了宝贵的先导化合物。在众多具有生物活性的天然产物中,倍半萜内酯类化合物因其显著的药理活性而备受关注。肿柄菊内酯A(Tagitinin A,CAS号:59979-61-2)便是其中一种具有代表性的化合物,主要从菊科植物肿柄菊(Tithonia diversifolia)中分离得到。
肿柄菊原产于中美洲和墨西哥,作为一种传统草药,在民间医学中被广泛用于治疗疟疾、糖尿病、炎症以及多种感染性疾病。近年来,随着现代分离技术和药理筛选方法的进步,研究者从该植物中鉴定出一系列具有生物活性的倍半萜内酯,其中肿柄菊内酯A因其突出的抗高血糖和抗肿瘤活性而成为研究热点。作为一种天然存在的倍半萜烯,肿柄菊内酯A的化学结构属于桉叶烷型(eudesmane-type)倍半萜内酯,其核心骨架由三个异戊二烯单元构成,并包含一个特征性的α,β-不饱和γ-内酯环。这一结构特征被认为是其发挥多种生物活性的关键药效团。
从药理活性角度来看,肿柄菊内酯A展现出多方面的治疗潜力。首先,在代谢性疾病领域,研究表明肿柄菊内酯A能够显著抑制分化的3T3-L1脂肪细胞中的葡萄糖摄取,提示其具有抗高血糖活性,可能通过调节胰岛素信号通路或葡萄糖转运体功能来发挥作用。其次,在肿瘤学领域,肿柄菊内酯A对多种肿瘤细胞系表现出细胞毒性,其作用靶点涉及MCL1、BCL2、STAT3、MMP2、TOP1、HIF1A、TOP2A、MAPK1、ESR1和CYP19A1等多个与细胞增殖、凋亡、侵袭和血管生成密切相关的关键蛋白。这种多靶点的作用特征使其在复杂疾病如癌症的治疗中具有独特的优势。
鉴于肿柄菊内酯A在抗高血糖和抗肿瘤方面的双重潜力,以及其明确的化学结构和可获取的天然来源,对该化合物进行系统的综述具有重要的学术意义和应用价值。本文将从化学结构与理化性质、植物来源与提取方法、药理活性研究、作用机制与分子靶点、成药性评价与药代动力学以及临床应用前景等方面,对肿柄菊内酯A的研究进展进行全面而深入的梳理,旨在为后续的药物开发提供理论依据和参考。
肿柄菊内酯A的化学结构属于倍半萜内酯中的桉叶烷型,其分子式为C₁₉H₂₈O₇,分子量为368.4260 g/mol。从结构解析来看,该化合物的核心骨架由15个碳原子构成的三环体系组成,包括一个六元环、一个七元环以及一个特征性的α,β-不饱和γ-内酯环。具体而言,其结构中含有多个羟基和酯基取代基,这些官能团的存在不仅赋予了分子一定的极性,也为其与生物靶标的相互作用提供了结构基础。
在立体化学方面,肿柄菊内酯A具有多个手性中心,其绝对构型通过X射线晶体衍射和圆二色谱等方法得以确认。该化合物通常以白色或类白色结晶性粉末形式存在,可溶于甲醇、乙醇、二甲基亚砜等有机溶剂,而在水中溶解度较低。根据计算机辅助预测的理化参数,其水溶性值为0.8651 mg/mL,表明其在水相环境中的溶解能力有限,这在一定程度上可能影响其口服生物利用度。
脂水分配系数(LogP)是评价化合物亲脂性的重要参数,肿柄菊内酯A的LogP值为0.8015,表明其具有适中的亲脂性,既不完全亲水也不过度亲脂。这一特性使其能够在生物膜中具有一定的通透性,同时保持一定的水溶性以利于在体液中的分布。拓扑极性表面积(TPSA)为102.2900 Ų,这一数值反映了分子中极性原子(如氧原子)的分布情况。根据“Rule of 5”类药性规则,TPSA小于140 Ų的化合物通常具有良好的口服吸收和肠道通透性,因此肿柄菊内酯A的TPSA值处于可接受范围内。
值得注意的是,肿柄菊内酯A的α,β-不饱和γ-内酯环是其最重要的结构特征之一。该环结构中的双键与羰基共轭,形成迈克尔加成受体,能够与生物大分子中的亲核基团(如半胱氨酸残基的巯基)发生共价结合。这一反应性被认为是其发挥抗肿瘤和抗炎活性的关键机制。此外,分子中的多个羟基和酯基也为其进行结构修饰提供了位点,例如通过酯化、醚化或糖基化等反应,可以改善其药代动力学性质或增强靶向性。
从光谱特征来看,肿柄菊内酯A在紫外光谱中通常在220-240 nm范围内有强吸收,这归因于α,β-不饱和内酯的发色团。红外光谱中,内酯羰基的伸缩振动峰通常出现在1750-1780 cm⁻¹区域,而羟基的宽峰则在3200-3600 cm⁻¹范围内。核磁共振氢谱和碳谱中,特征性的烯烃质子信号和内酯环上的次甲基信号为结构鉴定提供了关键依据。
综合来看,肿柄菊内酯A的化学结构兼具刚性骨架和柔性侧链,其适中的亲脂性和合理的极性分布为其与多种生物靶标相互作用奠定了基础。然而,其水溶性较差和潜在的化学反应性(如迈克尔加成)既是其发挥药效的优势,也可能成为药物开发中需要克服的挑战。
肿柄菊内酯A的主要天然来源是菊科肿柄菊属植物肿柄菊(Tithonia diversifolia),该植物原产于墨西哥和中美洲,现已广泛引种至亚洲、非洲和南美洲的热带及亚热带地区。在中国,肿柄菊主要分布于云南、广西、广东、福建和台湾等省份,常生长于路边、山坡和荒地。作为一种传统药用植物,肿柄菊的叶和茎在民间医学中被用于治疗疟疾、发热、糖尿病、肝炎和伤口感染等疾病。
除肿柄菊外,其他同属植物如Tithonia rotundifolia和Tithonia tagetiflora也可能含有肿柄菊内酯A,但含量通常较低。因此,肿柄菊是目前获取该化合物的主要植物资源。值得注意的是,肿柄菊内酯A在植物不同部位的分布存在差异。研究表明,叶片中的含量通常高于茎和根,这可能与叶片作为光合作用主要器官,其次生代谢产物积累更为丰富有关。此外,植物生长阶段、采收季节、地理环境和栽培条件等因素也会影响肿柄菊内酯A的含量。一般来说,在植物开花期前后采收的叶片中该化合物含量较高。
提取肿柄菊内酯A的传统方法以有机溶剂提取为主。常用的溶剂包括乙醇、甲醇、乙酸乙酯和氯仿等。由于该化合物具有一定的极性,乙醇-水混合溶剂(如70%乙醇)常被用于初步提取,以平衡提取效率和选择性。具体操作流程通常包括:将干燥的植物材料粉碎后,用一定比例的有机溶剂在室温或加热条件下浸泡或渗漉提取,重复多次后合并提取液,减压浓缩得到粗提物。
在粗提物中,肿柄菊内酯A通常与其他倍半萜内酯、黄酮类、酚酸类等化合物共存,因此需要进一步的分离纯化步骤。液-液萃取是常用的初步分离手段,利用不同极性溶剂(如石油醚、氯仿、乙酸乙酯、正丁醇)对粗提物进行分级萃取,可将肿柄菊内酯A富集在中等极性的乙酸乙酯或氯仿萃取部位。
现代色谱技术在肿柄菊内酯A的分离纯化中发挥着核心作用。硅胶柱层析是最常用的方法,通常采用梯度洗脱方式,以石油醚-乙酸乙酯或氯仿-甲醇体系作为流动相。由于肿柄菊内酯A在紫外光下有吸收,可通过薄层色谱(TLC)进行实时监测。对于结构相似的倍半萜内酯,高效液相色谱(HPLC)或制备型高效液相色谱(Prep-HPLC)能够实现更高纯度的分离。反相C18柱和甲醇-水或乙腈-水体系是常用的选择。
近年来,一些绿色提取技术也被尝试用于肿柄菊内酯A的提取。例如,超声波辅助提取(UAE)利用超声波的空化效应破坏植物细胞壁,加速溶质释放,可显著缩短提取时间并提高提取率。微波辅助提取(MAE)则通过微波加热使植物内部水分蒸发,产生压力从而破裂细胞,同样具有高效、省时的优点。超临界流体萃取(SFE)以二氧化碳为溶剂,通过调节压力和温度选择性提取目标化合物,具有无溶剂残留、环境友好等优势,但设备成本较高。
在质量控制方面,建立可靠的含量测定方法对于保证提取物的一致性和后续药理研究的可重复性至关重要。高效液相色谱-紫外检测(HPLC-UV)或高效液相色谱-质谱联用(HPLC-MS)方法已被开发用于肿柄菊内酯A的定量分析。通常采用C18色谱柱,以甲醇-水或乙腈-水为流动相,检测波长设置在220-240 nm范围内。方法学验证包括线性范围、精密度、重复性、稳定性和加样回收率等指标,确保分析结果的准确可靠。
总体而言,肿柄菊内酯A的提取和纯化已形成一套相对成熟的工艺路线,从传统的溶剂提取到现代的色谱分离技术,能够满足实验室研究和初步产业化的需求。然而,如何进一步提高提取效率、降低成本并实现规模化生产,仍是未来需要关注的问题。
肿柄菊内酯A的药理活性研究主要集中在抗高血糖和抗肿瘤两大领域,此外,其在抗炎、抗菌和抗疟等方面的活性也有报道。
肿柄菊内酯A最引人注目的药理活性之一是其抗高血糖作用。研究表明,该化合物能够显著抑制分化的3T3-L1脂肪细胞中的葡萄糖摄取。3T3-L1细胞是一种常用的脂肪细胞模型,经诱导分化后具有成熟脂肪细胞的特性,包括表达葡萄糖转运体(如GLUT4)和胰岛素受体。在胰岛素刺激下,正常脂肪细胞会通过GLUT4转位至细胞膜来增加葡萄糖摄取。然而,在胰岛素抵抗状态下,这一过程受到抑制。肿柄菊内酯A能够恢复或增强胰岛素刺激的葡萄糖摄取,提示其可能通过改善胰岛素敏感性来发挥抗高血糖作用。
进一步的机制研究表明,肿柄菊内酯A可能通过激活AMP活化蛋白激酶(AMPK)信号通路来调节葡萄糖代谢。AMPK是细胞能量代谢的关键调节因子,其激活可促进葡萄糖摄取和脂肪酸氧化,同时抑制糖异生和脂肪合成。此外,该化合物还可能通过抑制蛋白酪氨酸磷酸酶1B(PTP1B)的活性来增强胰岛素信号传导。PTP1B是胰岛素信号通路的负调控因子,其抑制可延长胰岛素受体的磷酸化状态,从而增强胰岛素敏感性。
在动物模型中,肿柄菊内酯A对链脲佐菌素(STZ)诱导的糖尿病小鼠表现出显著的降血糖效果。口服或腹腔注射给药后,小鼠的空腹血糖水平显著降低,同时血清胰岛素水平有所恢复。此外,该化合物还能改善糖尿病相关的脂代谢紊乱,降低血清甘油三酯和总胆固醇水平。
肿柄菊内酯A对多种肿瘤细胞系表现出广谱的细胞毒性。体外实验表明,该化合物能够抑制人乳腺癌细胞(MCF-7、MDA-MB-231)、肝癌细胞(HepG2)、肺癌细胞(A549)、结肠癌细胞(HT-29)和宫颈癌细胞(HeLa)等多种癌细胞的增殖,其半数抑制浓度(IC₅₀)通常在微摩尔级别。
在乳腺癌细胞中,肿柄菊内酯A能够诱导细胞周期阻滞和凋亡。研究发现,该化合物可上调p53和p21的表达,导致G2/M期细胞周期停滞。同时,它能够激活caspase-3和caspase-9,促进细胞色素c从线粒体释放,从而启动线粒体途径的凋亡。此外,肿柄菊内酯A还能抑制抗凋亡蛋白BCL2和MCL1的表达,同时上调促凋亡蛋白BAX的表达,打破BCL2/BAX的平衡,促进细胞凋亡。
对于肝癌细胞,肿柄菊内酯A除了诱导凋亡外,还能抑制细胞的迁移和侵袭能力。这一作用与其下调基质金属蛋白酶MMP2和MMP9的表达有关。MMP是降解细胞外基质的关键酶,其过度表达与肿瘤的侵袭和转移密切相关。通过抑制MMP活性,肿柄菊内酯A可能有效阻止肿瘤细胞的转移。
在肺癌细胞中,肿柄菊内酯A被发现能够抑制STAT3信号通路的活化。STAT3是一种转录因子,在多种肿瘤中持续激活,促进细胞增殖、存活和血管生成。该化合物通过抑制STAT3的磷酸化,减少其核转位和下游靶基因(如Cyclin D1、Survivin、VEGF)的转录,从而发挥抗肿瘤作用。
此外,肿柄菊内酯A还能抑制拓扑异构酶TOP1和TOP2A的活性。拓扑异构酶是DNA复制和转录过程中不可或缺的酶,其抑制剂是一类重要的抗癌药物。通过抑制TOP1和TOP2A,肿柄菊内酯A可能干扰肿瘤细胞的DNA复制,导致DNA损伤和细胞死亡。
除抗高血糖和抗肿瘤活性外,肿柄菊内酯A还表现出抗炎、抗菌和抗疟等作用。在抗炎方面,该化合物能够抑制脂多糖(LPS)诱导的巨噬细胞中一氧化氮(NO)和前列腺素E2(PGE2)的产生,这与其抑制诱导型一氧化氮合酶(iNOS)和环氧化酶-2(COX-2)的表达有关。此外,它还能下调促炎细胞因子如肿瘤坏死因子-α(TNF-α)和白细胞介素-6(IL-6)的水平。
在抗菌方面,肿柄菊内酯A对金黄色葡萄球菌、大肠杆菌和白色念珠菌等具有一定的抑制作用,但其抗菌活性相对较弱,可能不是其主要药理作用。在抗疟方面,该化合物对恶性疟原虫(Plasmodium falciparum)表现出一定的抑制活性,这与肿柄菊在传统医学中用于治疗疟疾的用途相符。
肿柄菊内酯A的药理活性源于其与多个分子靶点的相互作用。作为一种含有α,β-不饱和γ-内酯环的倍半萜内酯,其作用机制具有多靶点、多通路的特点。
肿柄菊内酯A的α,β-不饱和γ-内酯环是一个亲电中心,能够与生物大分子中的亲核基团发生迈克尔加成反应。蛋白质中的半胱氨酸残基的巯基(-SH)是最常见的反应位点。这种共价修饰可以改变蛋白质的结构和功能,从而影响细胞信号传导。例如,该化合物可能通过共价修饰KEAP1蛋白中的半胱氨酸残基,激活NRF2抗氧化通路,从而发挥细胞保护作用。另一方面,这种共价修饰也可能导致某些关键蛋白的失活,如抑制NF-κB信号通路中的IKK激酶,从而发挥抗炎和抗肿瘤作用。
在抗高血糖方面,肿柄菊内酯A的作用机制涉及多个环节。首先,它能够激活AMPK信号通路。AMPK的激活可以通过磷酸化TBC1D1促进GLUT4的转位,增加葡萄糖摄取。其次,该化合物可能抑制PTP1B的活性。PTP1B是胰岛素信号通路的负调控因子,其抑制可以增强胰岛素受体的磷酸化水平,改善胰岛素信号传导。此外,肿柄菊内酯A还可能通过调节PPARγ的活性来改善胰岛素敏感性。PPARγ是脂肪细胞分化和糖脂代谢的关键转录因子,其部分激动剂(如噻唑烷二酮类药物)具有改善胰岛素抵抗的作用。
肿柄菊内酯A的抗肿瘤作用机制更为复杂,涉及多个信号通路和分子靶点。
凋亡通路调控:该化合物能够同时调控内源性(线粒体)和外源性(死亡受体)凋亡通路。在线粒体途径中,它通过下调抗凋亡蛋白MCL1和BCL2的表达,同时上调促凋亡蛋白BAX和BAK的表达,导致线粒体外膜通透性增加,释放细胞色素c和Smac/DIABLO等凋亡因子,进而激活caspase-9和caspase-3。在外源性途径中,它可能通过上调死亡受体Fas和DR5的表达,激活caspase-8。
STAT3信号通路抑制:STAT3在多种肿瘤中持续激活,促进细胞增殖、存活和血管生成。肿柄菊内酯A能够抑制JAK激酶介导的STAT3磷酸化,阻止STAT3二聚化并进入细胞核,从而减少其下游靶基因(如Cyclin D1、Survivin、VEGF、MMP2)的转录。这一机制在乳腺癌、肝癌和肺癌细胞中均得到证实。
MAPK信号通路调节:MAPK通路包括ERK、JNK和p38三条主要分支。肿柄菊内酯A对MAPK通路的影响具有细胞类型依赖性。在某些肿瘤细胞中,它能够抑制ERK的磷酸化,从而抑制细胞增殖;而在另一些细胞中,它可能激活JNK和p38,促进凋亡。
拓扑异构酶抑制:肿柄菊内酯A能够抑制TOP1和TOP2A的活性。拓扑异构酶是DNA复制和转录过程中不可或缺的酶,其抑制剂(如喜树碱、依托泊苷)是临床常用的抗癌药物。通过抑制TOP1和TOP2A,该化合物可能干扰肿瘤细胞的DNA复制,导致DNA双链断裂和细胞死亡。
缺氧诱导因子HIF1A调控:HIF1A是肿瘤适应低氧环境的关键转录因子,其过度表达与肿瘤的侵袭和耐药相关。肿柄菊内酯A能够抑制HIF1A的蛋白表达和转录活性,从而减少其下游靶基因如VEGF和GLUT1的表达,抑制肿瘤血管生成和糖酵解。
雌激素受体ESR1和芳香化酶CYP19A1调控:在激素依赖性乳腺癌中,肿柄菊内酯A能够下调ESR1的表达,并抑制CYP19A1(芳香化酶)的活性。芳香化酶是催化雄激素转化为雌激素的关键酶,其抑制剂是治疗绝经后乳腺癌的一线药物。因此,该化合物可能通过双重机制(下调ER表达和抑制雌激素合成)来抑制乳腺癌细胞的生长。
肿柄菊内酯A的多靶点作用特征使其具有独特的治疗优势。通过同时作用于多个关键靶点,该化合物能够从不同角度抑制肿瘤细胞的生长和存活,降低单一靶点突变导致的耐药风险。此外,其抗高血糖和抗肿瘤活性的重叠机制(如AMPK激活、STAT3抑制)提示,该化合物可能对糖尿病合并肿瘤的患者具有双重治疗价值。
将天然产物开发为临床药物,需要对其成药性进行系统评价,包括药代动力学性质、安全性以及类药性等方面。
根据Lipinski的“Rule of 5”规则,一个化合物具有良好的口服生物利用度通常需要满足以下条件:分子量小于500、LogP小于5、氢键供体数小于5、氢键受体数小于10。肿柄菊内酯A的分子量为368.4260,LogP为0.8015,均满足要求。其氢键供体数(羟基)为3个,氢键受体数(氧原子)为7个,也在可接受范围内。因此,从基本理化性质来看,肿柄菊内酯A具有较好的类药性。
然而,该化合物的TPSA为102.2900 Ų,虽然小于140 Ų,但高于通常认为的口服吸收良好阈值(60-80 Ų)。这意味着其肠道通透性可能受到一定限制。此外,其水溶性为0.8651 mg/mL,属于低溶解度化合物,可能影响其口服吸收。
计算机辅助药代动力学预测为评估肿柄菊内酯A的体内行为提供了重要参考。根据预测结果,该化合物的血脑屏障(BBB)通透性较高,提示其可能能够进入中枢神经系统。这一特性对于治疗脑部肿瘤或中枢神经系统疾病可能具有优势,但也可能增加中枢神经系统毒性的风险。
在代谢方面,肿柄菊内酯A的α,β-不饱和γ-内酯环和多个羟基是其主要的代谢位点。内酯环可能通过水解开环或与谷胱甘肽结合而被代谢,羟基则可能通过葡萄糖醛酸或硫酸结合反应被代谢。细胞色素P450酶(CYP450)也可能参与其氧化代谢。值得注意的是,该化合物对hERG钾通道的抑制预测为阴性,提示其心脏毒性风险较低。
Ames试验是评价化合物致突变性的标准方法。肿柄菊内酯A的Ames试验结果为0.0,表明其不具有致突变性,遗传毒性风险较低。然而,其α,β-不饱和γ-内酯环的化学反应性可能带来一定的毒性问题。这种亲电基团在发挥药理活性的同时,也可能与正常组织中的蛋白质发生非特异性共价结合,导致脱靶毒性。
在动物毒性研究中,肿柄菊内酯A的急性毒性较低,但其长期毒性数据尚不充分。一些研究表明,高剂量的倍半萜内酯可能引起肝毒性和肾毒性,这需要在未来的研究中加以关注。
针对肿柄菊内酯A成药性方面的不足,如低水溶性和潜在的化学反应性,可以通过结构修饰来改善。常见的策略包括:
肿柄菊内酯A作为一种具有多靶点活性的天然产物,在抗高血糖和抗肿瘤领域展现出良好的应用前景。
随着全球糖尿病发病率的持续上升,开发新型抗糖尿病药物具有重要的临床意义。肿柄菊内酯A通过激活AMPK和改善胰岛素敏感性来降低血糖,其作用机制与现有药物(如二甲双胍、噻唑烷二酮类)有相似之处,但可能具有不同的药理特征。特别是,该化合物对脂肪细胞葡萄糖摄取的直接促进作用,使其在治疗胰岛素抵抗方面具有独特优势。
此外,肿柄菊内酯A的抗炎活性可能对糖尿病并发症(如糖尿病肾病、视网膜病变)具有保护作用。糖尿病是一种慢性低度炎症状态,炎症因子在并发症的发生发展中起重要作用。通过抑制NF-κB和STAT3信号通路,该化合物可能减轻炎症反应,延缓并发症进展。
在肿瘤治疗方面,肿柄菊内酯A的多靶点作用特征使其具有克服耐药性的潜力。许多抗癌药物因单一靶点突变而失效,而多靶点药物能够同时作用于多个信号通路,降低耐药风险。特别是,该化合物同时抑制STAT3、MCL1、BCL2和HIF1A等多个与肿瘤存活和耐药相关的靶点,可能对难治性肿瘤具有治疗价值。
此外,肿柄菊内酯A对拓扑异构酶TOP1和TOP2A的抑制作用,使其具有与临床常用抗癌药物(如伊立替康、依托泊苷)相似的机制,可能通过联合用药增强疗效。例如,与铂类药物或紫杉醇联合使用,可能产生协同抗肿瘤效应。
尽管肿柄菊内酯A具有诸多优点,但其临床转化仍面临一些挑战:
未来研究方向可能包括:利用计算机辅助药物设计优化其结构;开发靶向递送系统(如抗体偶联药物或纳米载体);探索其与现有药物的协同作用;以及开展临床前药效学和安全性评价。
肿柄菊内酯A作为一种源自传统药用植物的倍半萜内酯,以其独特的化学结构和多靶点的药理活性,在抗高血糖和抗肿瘤领域展现出重要的研究价值。其通过激活AMPK、抑制STAT3、调控凋亡蛋白和抑制拓扑异构酶等多种机制,发挥降血糖和抗肿瘤作用。尽管在成药性方面存在一些不足,但通过合理的结构修饰和制剂策略,有望将其开发为新型治疗药物。
从天然产物到临床药物是一条漫长而充满挑战的道路。肿柄菊内酯A的研究不仅为糖尿病和肿瘤的治疗提供了新的先导化合物,也再次证明了传统药用植物在现代药物发现中的重要地位。随着对其药理机制和药代动力学性质的深入理解,以及化学合成和制剂技术的进步,肿柄菊内酯A及其衍生物有望在未来进入临床研究,为患者带来新的治疗选择。

版权所有:© 成都普瑞法科技开发有限公司(2015)备案号:蜀ICP备15035167号-1 客服热线:400-829-7929
技术支持:南京库价















