
产品名称: 三七皂苷R3
英文名: Notoginsenoside R3
中文别名:
英文别名:
Cas 号: 87741-76-2
产品编码:BP5290
分子式: C48H82O19
分子量: 963.165
来源:
化合物类型: 三萜类(Triterpenoids)
纯度: 95%~99%
分析方法: HPLC-DAD or/and HPLC-ELSD
鉴定方法: 质谱(Mass), 核磁(NMR)
包装: 棕色小玻璃瓶,按客户需求包装。
存储: 贮存在避光密闭容器中,冷藏或者冷冻长期保存。
样品溶液最好临用新配。如果需要提前配制的话,最好分成独立包装冷冻保存(-20℃以下),临用前再取出解冻,通常可以保存2周。
可以满足克级以上大量需求,详情请咨询。
提供相关图谱和分析方法指导,可以提供包括色谱柱,标准品和分析方法在内的含量测定整体服务包,价格优惠。
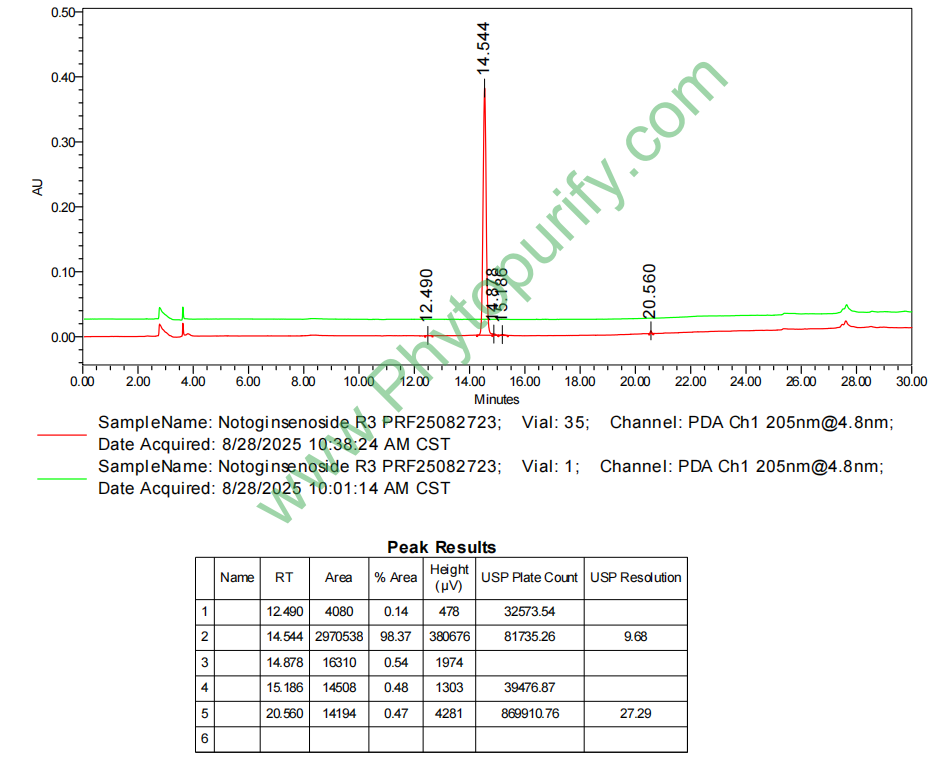
三七皂苷R3的HPLC图谱
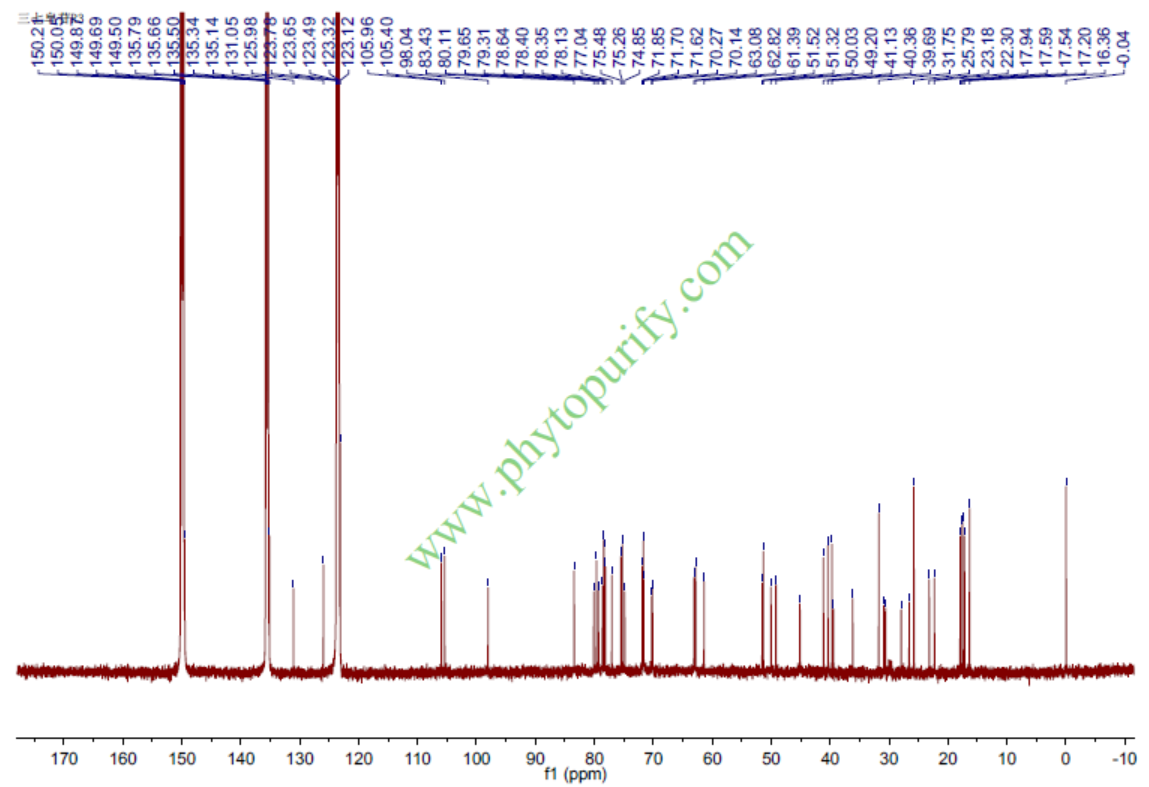
三七皂苷R3的核磁图谱
储存条件:短期保存 2~8℃,长期保存 -20 ~ -80℃
318.37
1.99
1.99
0.15
0.47
0.16
低
63.93
6.52
否
否
否
否
无
无
0.0
是
否
否
否

三七皂苷R3(Notoginsenoside R3,简称NGR3)是从五加科人参属植物三七(Panax notoginseng (Burk.) F.H. Chen)中分离得到的一种达玛烷型三萜皂苷。三七作为传统名贵中药,在我国已有数百年的临床应用历史,其根茎常被用于活血化瘀、消肿定痛,尤其在心血管疾病的防治中占据重要地位。现代药理学研究表明,三七的主要活性成分包括三七皂苷(Notoginsenosides)和人参皂苷(Ginsenosides),其中三七皂苷R3因其独特的化学结构和显著的生物活性,近年来逐渐成为天然产物研究领域的热点。
与三七中含量较高的主要皂苷(如人参皂苷Rb1、Rg1)不同,三七皂苷R3属于微量皂苷,但其生物活性却不容小觑。早期研究多聚焦于三七总皂苷对心血管系统的保护作用,而随着分离纯化技术的进步和活性导向筛选策略的应用,NGR3的独特药理价值得以揭示。现有证据表明,NGR3在免疫调节、抗炎、抗肿瘤以及组织保护等方面展现出多效性,其作用机制涉及对多条信号通路的精细调控,尤其是对免疫稳态的维持具有重要影响。本文旨在系统梳理三七皂苷R3的化学特性、植物来源、药理活性、分子机制及成药性,以期为该化合物的深入开发与临床转化提供理论依据。
三七皂苷R3的化学结构属于达玛烷型四环三萜皂苷,其母核为20(S)-原人参二醇(Protopanaxadiol, PPD)。与常见的原人参二醇型皂苷(如人参皂苷Rb1)相比,NGR3的糖链取代模式具有独特性。具体而言,其糖基部分由葡萄糖和木糖等单糖组成,通常连接于C-3和C-20位羟基上。根据现有文献报道,NGR3的完整化学名为:3-O-β-D-吡喃葡萄糖基-(1→2)-β-D-吡喃葡萄糖基-20-O-β-D-吡喃木糖基-(1→6)-β-D-吡喃葡萄糖基-20(S)-原人参二醇。其分子式为C₄₇H₈₀O₂₀,分子量精确值为963.1650 Da。
从理化性质来看,NGR3表现出典型的三萜皂苷特征。其脂水分配系数(LogP)为1.9885,表明该化合物具有一定的亲脂性,但总体偏向于中等极性。拓扑极性表面积(TPSA)高达318.3700 Ų,这主要归因于其分子中大量的羟基和糖苷键,使其具有较强的形成氢键的能力。水溶性参数为0.1534 mg/mL,属于微溶范畴,这在一定程度上限制了其口服生物利用度。值得注意的是,NGR3的分子量较大(>900 Da),且TPSA极高,根据“Lipinski五规则”及“Veber规则”,该化合物在口服吸收方面可能面临挑战。此外,计算机模拟预测显示,NGR3穿透血脑屏障(BBB)的能力较低,这提示其药理作用主要集中于外周组织。在安全性方面,hERG抑制预测结果为阴性,Ames试验结果为0.0,表明该化合物在基因毒性和心脏毒性方面具有较低的潜在风险,为其后续开发提供了有利的安全性窗口。
三七皂苷R3主要来源于五加科植物三七(Panax notoginseng)的干燥根及根茎。三七主要分布于中国云南文山、广西百色等地,其生长周期较长,通常需三年以上方可采收。NGR3在三七中的含量相对较低,属于微量成分,远低于人参皂苷Rg1、Rb1及三七皂苷R1等主要皂苷。因此,其获取往往依赖于高效的提取与分离技术。
传统的提取方法多采用乙醇或甲醇回流提取,随后通过正丁醇萃取富集总皂苷。然而,由于NGR3与结构类似物(如人参皂苷Rd、Rb1等)的极性相近,常规的硅胶柱层析难以实现有效分离。近年来,随着现代色谱技术的发展,高速逆流色谱(HSCCC)、制备型高效液相色谱(Pre-HPLC)以及大孔吸附树脂联用技术被广泛应用于NGR3的分离纯化。例如,采用D101型大孔吸附树脂对三七总皂苷进行初步分级,再结合ODS反相柱层析和制备型HPLC,可成功获得纯度大于98%的NGR3单体。此外,基于分子印迹技术或亲和色谱的新型分离方法也在探索之中,旨在提高微量皂苷的回收率。
值得注意的是,NGR3不仅存在于三七原植物中,也可能在人参属其他植物(如人参、西洋参)中微量存在,但以三七中的含量和生物活性最为突出。由于NGR3在植物体内含量极低,生物合成途径的研究也逐步展开。通过转录组学分析,已初步鉴定出参与NGR3糖基化修饰的关键糖基转移酶基因,这为未来通过合成生物学手段异源生产NGR3提供了可能。
免疫调节是三七皂苷R3最受关注的药理活性之一。现有研究表明,NGR3能够双向调节免疫应答,即在抑制过度炎症反应的同时,增强机体对病原体的清除能力。在体外实验中,NGR3可显著调节巨噬细胞和树突状细胞的功能。例如,在脂多糖(LPS)刺激的RAW264.7巨噬细胞模型中,NGR3能够抑制促炎因子如肿瘤坏死因子-α(TNF-α)、白细胞介素-6(IL-6)和一氧化氮(NO)的释放,同时上调抗炎因子白细胞介素-10(IL-10)的表达。这种调节作用并非单纯的免疫抑制,而是通过重塑免疫微环境来实现稳态。
在适应性免疫方面,NGR3对T细胞的分化具有调控作用。研究发现,NGR3能够促进调节性T细胞(Treg)的分化,同时抑制Th17细胞的过度活化。这一效应与转录因子FOXP3和STAT3的表达变化密切相关。此外,NGR3还能影响B细胞的抗体分泌功能,提示其在体液免疫中也扮演重要角色。
除免疫调节外,NGR3展现出显著的抗炎和抗氧化活性。在多种炎症模型中,如角叉菜胶诱导的足趾肿胀模型和醋酸诱导的腹腔毛细血管通透性增加模型,NGR3均能有效减轻炎症反应。其机制与抑制核因子κB(NF-κB)的活化有关,NF-κB是炎症反应的核心转录因子,NGR3通过阻断IκBα的磷酸化降解,从而抑制p65亚基的核转位,最终减少下游炎症介质的转录。
在氧化应激方面,NGR3能够清除自由基,提高细胞内超氧化物歧化酶(SOD)和谷胱甘肽过氧化物酶(GSH-Px)的活性,降低丙二醛(MDA)水平。这种抗氧化保护作用在心肌细胞和肝细胞损伤模型中尤为明显。
鉴于三七在心血管疾病中的传统应用,NGR3的心血管保护作用也得到了初步验证。在心肌缺血再灌注损伤模型中,NGR3预处理可显著减少心肌梗死面积,降低乳酸脱氢酶(LDH)和肌酸激酶(CK)的释放。其保护机制涉及激活PI3K/Akt信号通路,抑制线粒体通透性转换孔(mPTP)的开放,从而减少细胞凋亡。此外,NGR3还能抑制血小板聚集,延长凝血时间,提示其具有抗血栓形成的潜力。
近年来,NGR3的抗肿瘤活性逐渐引起关注。在多种肿瘤细胞系(如肝癌HepG2、肺癌A549、乳腺癌MCF-7)中,NGR3表现出剂量依赖性的增殖抑制效应。其作用机制包括诱导细胞周期阻滞于G0/G1期,以及通过线粒体途径和内质网应激途径诱导凋亡。值得注意的是,NGR3对正常细胞的毒性较低,显示出一定的选择性。此外,NGR3还能增强化疗药物(如顺铂、紫杉醇)的敏感性,可能通过抑制NF-κB介导的耐药机制实现。
三七皂苷R3的药理作用是多靶点、多通路协同作用的结果。基于现有研究,其核心分子机制可归纳为以下几个方面:
Toll样受体4(TLR4)是识别LPS等病原相关分子模式的关键受体。NGR3能够直接结合TLR4的胞外域,竞争性抑制LPS与TLR4/MD2复合物的结合,从而阻断下游MyD88依赖性和TRIF依赖性信号转导。这导致IκB激酶(IKK)复合物的活化受阻,NF-κB(由NFKB1编码的p50和RelA组成)无法进入细胞核启动促炎基因(如IL2、IFNG、TNF)的转录。这一机制是NGR3发挥抗炎和免疫抑制效应的基础。
信号转导及转录激活因子(STAT)家族在免疫细胞分化中起决定性作用。NGR3可调节STAT3和STAT4的磷酸化水平。在T细胞中,NGR3通过抑制STAT3的活化,减少Th17细胞的分化;同时,通过促进STAT5的磷酸化,增强Treg细胞的发育。此外,NGR3还能上调细胞因子信号转导抑制因子(SOCS)的表达,形成负反馈调节环,从而精细调控免疫应答的强度。
转化生长因子-β1(TGFB1)是诱导Treg细胞分化的关键因子。NGR3能够上调TGF-β1的表达,并激活下游Smad2/3信号,协同促进FOXP3的表达。同时,NGR3可增强细胞毒性T淋巴细胞相关蛋白4(CTLA4)的表达,CTLA4作为免疫检查点分子,能够抑制T细胞的过度活化,维持免疫耐受。这一机制解释了NGR3在自身免疫性疾病模型中的保护作用。
综合来看,NGR3的分子靶点网络以TLR4、STAT3、NFKB1、TGFB1、CTLA4、FOXP3、IL10和IFNG为核心节点。其中,IL10作为抗炎因子,其表达上调是NGR3发挥免疫调节作用的重要效应环节;而IFNG(干扰素-γ)的表达则受到精细调控,在抗感染免疫中保持适度水平。这种多靶点的协同调控模式,使得NGR3能够在不完全抑制免疫系统的情况下,纠正免疫失衡状态。
从药物化学角度,NGR3的成药性面临一定挑战。其分子量(963.17 Da)远超口服药物的常规阈值(500 Da),且TPSA(318.37 Ų)极高,这导致其跨膜渗透性较差。根据“Lipinski五规则”,NGR3违反了分子量、氢键供体数和氢键受体数三条规则,属于典型的“beyond Rule of 5”(bRo5)化合物。然而,bRo5化合物并非完全不可成药,许多天然产物(如环孢素A)和大环内酯类药物均属于此类,其口服吸收往往依赖于主动转运或淋巴转运途径。
NGR3的LogP为1.99,处于适中范围,表明其亲脂性尚可,但水溶性(0.1534 mg/mL)偏低,可能影响制剂开发。安全性方面,hERG抑制和Ames试验均为阴性,提示其心脏毒性和遗传毒性风险较低,这是其成药性的重要优势。
目前关于NGR3药代动力学的体内研究尚不充分,但可参考结构类似物(如人参皂苷Rb1、Rd)的代谢特征进行推断。口服给药后,NGR3在胃肠道中可能经历酸水解和肠道菌群代谢,糖链逐步被切除,生成次级苷或苷元(如原人参二醇)。这些代谢产物可能具有不同的生物活性。由于分子量大且极性高,NGR3的口服生物利用度通常较低,可能低于5%。静脉给药后,NGR3在血浆中的分布半衰期较短,主要分布于肝脏、肾脏和肺组织,脑组织分布极少,与BBB预测结果一致。
代谢途径方面,NGR3主要经肝脏代谢,可能涉及去糖基化和氧化反应。排泄途径以胆汁排泄为主,部分代谢产物经粪便排出,尿液中原型药物含量极低。为提高其生物利用度,纳米脂质体、磷脂复合物、自微乳给药系统等新型递送技术正在被探索。
基于NGR3对TLR4/NF-κB和STAT3信号通路的调控作用,其在炎症性肠病、类风湿性关节炎、银屑病等自身免疫性疾病中具有潜在应用价值。特别是其能够促进Treg细胞分化并上调CTLA4表达的特性,使其可能成为一种新型的免疫耐受诱导剂。此外,NGR3在器官移植排斥反应和移植物抗宿主病(GVHD)中的保护作用也值得探索。
NGR3的双向免疫调节特性在肿瘤免疫治疗中展现出独特优势。一方面,它可以通过抑制NF-κB和STAT3来降低肿瘤微环境中的炎症水平,从而逆转免疫抑制状态;另一方面,它能够增强CTLA4的表达,这提示其可能与免疫检查点抑制剂(如抗CTLA-4抗体)产生协同或拮抗作用。未来的研究需要明确NGR3在肿瘤免疫周期中的具体作用节点。
作为三七的活性成分,NGR3在心血管疾病中的应用最为直接。其抗心肌缺血、抗血小板聚集和抗炎作用,使其有望成为治疗冠心病、心肌梗死和动脉粥样硬化的候选药物。然而,由于口服生物利用度低,开发注射剂型或经皮给药系统可能是更优的选择。
尽管前景广阔,NGR3的临床转化仍面临诸多挑战。首先,其植物来源含量极低,大规模生产依赖于化学合成或生物合成,成本较高。其次,药代动力学特性不佳,需要借助现代制剂技术改善其吸收和代谢稳定性。此外,目前关于NGR3的体内药效学研究多局限于动物模型,缺乏高质量的人体临床试验数据。未来,应结合系统药理学和网络药理学方法,进一步阐明NGR3的多靶点作用机制,并利用结构修饰策略(如前药设计、糖基化改造)优化其成药性。
三七皂苷R3作为三七中的一种微量活性成分,凭借其独特的达玛烷型结构和多效性药理活性,正在从传统中药的“配角”转变为现代药物研发的“新星”。本文系统梳理了其在化学结构、植物来源、药理活性、分子机制及成药性方面的研究进展,揭示了NGR3通过调控TLR4、STAT3、NF-κB、TGF-β1等关键靶点,发挥免疫调节、抗炎、心血管保护和抗肿瘤作用的分子基础。尽管在口服生物利用度和规模化制备方面仍存在瓶颈,但其良好的安全性窗口和独特的免疫稳态调节机制,使其在自身免疫性疾病、肿瘤免疫治疗及心血管疾病领域具有不可忽视的开发潜力。随着合成生物学、药物化学和纳米制剂技术的进步,三七皂苷R3有望从实验室走向临床,成为源自中药的创新型药物分子。

版权所有:© 成都普瑞法科技开发有限公司(2015)备案号:蜀ICP备15035167号-1 客服热线:400-829-7929
技术支持:南京库价















