
产品名称: 忍冬苷
英文名: Lonicerin
中文别名: 木犀草素-7-O-新橙皮糖苷
英文别名: Luteolin 7-neohesperidoside
Cas 号: 25694-72-8
产品编码:BP2007
分子式: C27H30O15
分子量: 594.522
来源:
物理性状:
化合物类型:
纯度: 95%~99%
分析方法: HPLC-DAD or/and HPLC-ELSD
鉴定方法: 质谱(Mass), 核磁(NMR)
包装: 棕色小玻璃瓶,标准包装10mg,20mg,50mg;可以按客户需求包装。
可以满足克级以上大量需求,详情请咨询。
提供相关图谱和分析方法指导,可以提供包括色谱柱,标准品和分析方法在内的含量测定整体服务包,价格优惠。
提供改善产品水溶性解决方案,改善化合物在水中的溶解度,便于进行各种活性试验和临床应用。
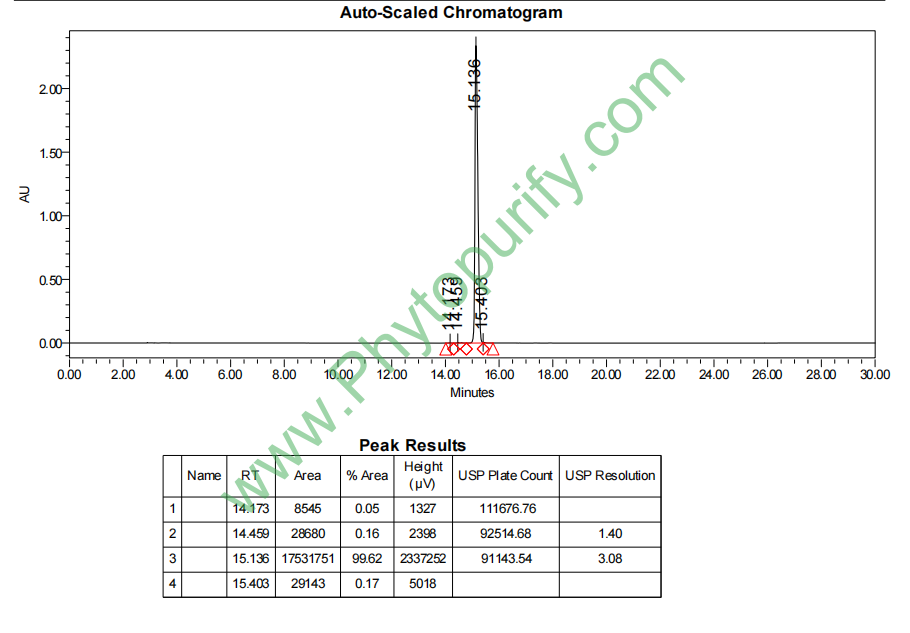
忍冬苷的HPLC图谱
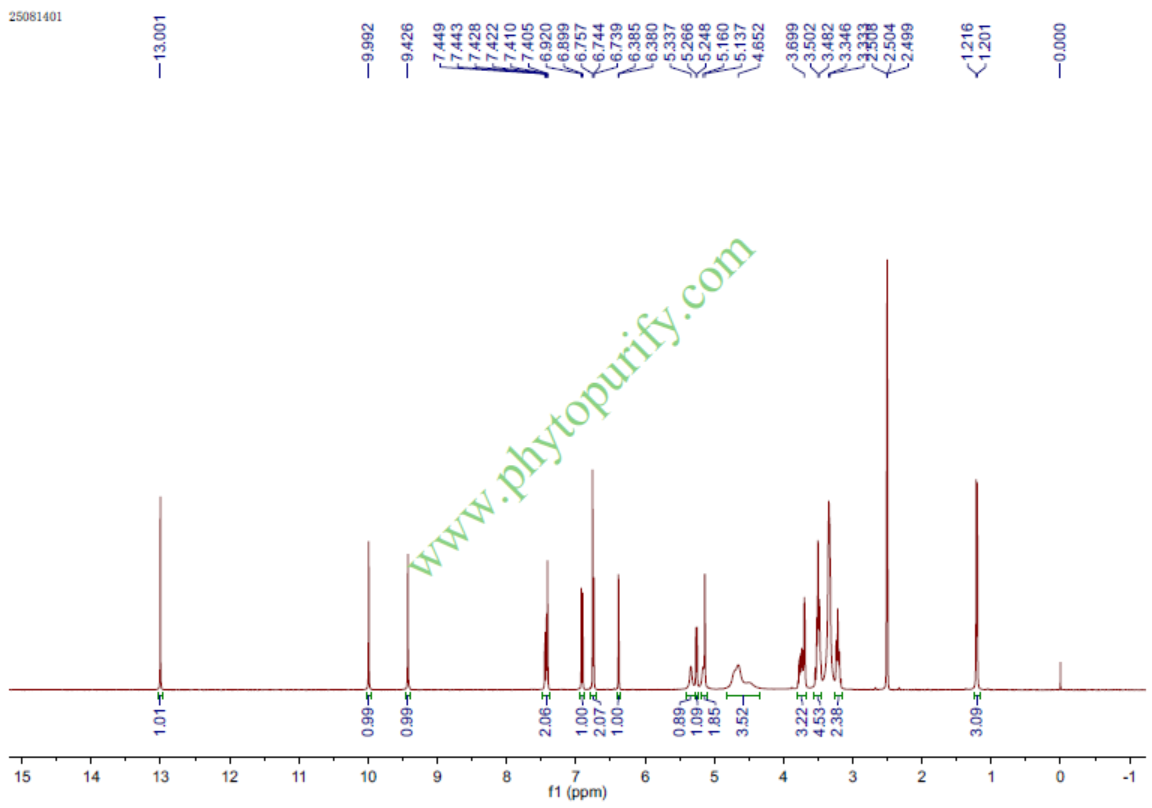
忍冬苷的核磁图谱
储存条件:短期保存 2~8℃,长期保存 -20 ~ -80℃
249.20
-0.22
-0.25
3.12
0.48
0.13
低
79.03
4.71
是
否
是
否
有
无
0.6
是
否
是
否

天然产物作为药物发现的重要源泉,在人类对抗疾病的漫长历史中扮演着不可替代的角色。黄酮类化合物作为植物次生代谢产物的重要组成部分,因其结构多样性和广泛的生物活性而备受关注。忍冬苷(Lonicerin),亦称Veronicastroside,是一种天然存在的黄酮苷类化合物,最早从忍冬科植物中分离鉴定。随着现代药理学研究的深入,忍冬苷展现出令人瞩目的多重生物活性,包括抗炎、抗氧化、抗关节炎、抗菌、抗真菌以及神经保护作用,尤其在抗流感病毒和调控相关信号通路方面显示出独特优势。
近年来,全球范围内流感病毒的持续变异和耐药性问题的日益严峻,使得开发新型抗流感药物成为紧迫的医学需求。传统抗流感药物如神经氨酸酶抑制剂(奥司他韦)和M2离子通道抑制剂(金刚烷胺)面临着耐药病毒株不断出现的困境。在此背景下,天然产物因其多靶点作用特点和较低的耐药性风险,成为抗流感药物研发的重要方向。忍冬苷通过调控NFE2L2、RELA、MAPK8、NFKB1等多个与流感病毒感染和炎症反应密切相关的靶点,展现出作为多靶点抗流感先导化合物的巨大潜力。
此外,忍冬苷对黄嘌呤氧化酶的抑制作用(IC50为37.4 µg/mL)提示其在痛风和高尿酸血症治疗中的应用前景;其对藻酸盐分泌蛋白AlgE的抑制活性则揭示了抗铜绿假单胞菌生物膜形成的潜在机制。这些发现共同勾勒出忍冬苷作为多功能天然药物的研究价值。本文将从化学结构、植物来源、药理活性、作用机制、成药性评价及临床应用前景等方面,对忍冬苷的研究进展进行系统综述。
忍冬苷的化学名为5,7-二羟基-2-(4-羟基苯基)-4H-1-苯并吡喃-4-酮-7-O-β-D-葡萄糖苷,属于黄酮苷类化合物。其苷元为木犀草素(Luteolin),糖基部分为葡萄糖。从结构解析来看,忍冬苷的母核为2-苯基色原酮结构,A环C-5和C-7位各有一个羟基,其中C-7位羟基与葡萄糖形成O-糖苷键。B环为4′-羟基苯基,形成典型的黄酮骨架。
忍冬苷的分子式为C27H30O15,分子量为594.5220 Da。该化合物含有多个酚羟基和糖基,赋予其良好的水溶性和抗氧化活性。从结构-活性关系角度分析,C-7位的糖基化修饰不仅影响化合物的溶解性和生物利用度,还可能改变其与靶蛋白的相互作用模式。与苷元木犀草素相比,忍冬苷的糖基部分可能降低其细胞膜通透性,但同时也可能增强其与某些细胞表面受体或转运蛋白的亲和力。
根据计算化学和实验测定结果,忍冬苷的主要理化性质参数如下:
这些理化性质参数为忍冬苷的药物开发提供了重要的参考依据。其良好的水溶性和安全性特征有利于制剂开发,但低的口服生物利用度和血脑屏障穿透性则是需要克服的挑战。
忍冬苷最初从忍冬科(Caprifoliaceae)植物中分离鉴定,因此得名。其主要植物来源包括:
忍冬(Lonicera japonica Thunb.):又名金银花,是忍冬苷最丰富的来源之一。金银花作为传统中药材,具有清热解毒、疏散风热的功效,常用于治疗感冒、发热、咽喉肿痛等症。忍冬苷被认为是金银花中重要的活性成分之一,与其抗炎、抗病毒作用密切相关。
水蔓菁(Veronica linariifolia Pall. ex Link):该植物是忍冬苷的另一个重要来源,化合物也因此得名Veronicastroside。水蔓菁在民间医学中用于治疗呼吸道感染和炎症性疾病。
其他来源:忍冬苷也存在于其他忍冬属植物(如Lonicera macranthoides、Lonicera confusa)以及部分玄参科植物中。不同植物来源的忍冬苷含量差异较大,通常以花蕾和叶片中含量较高。
忍冬苷的提取通常采用溶剂提取法,结合现代分离纯化技术,以获得高纯度的化合物。
提取工艺:
- 溶剂选择:由于忍冬苷具有较好的水溶性和醇溶性,常用提取溶剂包括水、甲醇、乙醇及其混合溶剂。研究表明,50%-70%乙醇水溶液对忍冬苷的提取效率较高。
- 提取方法:传统方法包括冷浸法、回流提取法和超声辅助提取法。超声辅助提取可显著缩短提取时间,提高提取效率。近年来,微波辅助提取和酶辅助提取等绿色提取技术也被应用于忍冬苷的提取,具有溶剂用量少、提取率高的优点。
- 提取条件优化:影响提取效率的关键因素包括溶剂浓度、料液比、提取温度、提取时间和提取次数。以金银花为例,优化的提取条件为:60%乙醇,料液比1:20,60℃超声提取30分钟,提取2次。
纯化方法:
- 大孔吸附树脂:是忍冬苷纯化最常用的方法之一。HPD-100、D101、AB-8等型号的大孔树脂对忍冬苷具有良好的吸附和解吸性能。通过梯度洗脱(通常使用乙醇-水体系),可获得纯度较高的忍冬苷粗品。
- 柱层析:硅胶柱层析、聚酰胺柱层析和葡聚糖凝胶柱层析(Sephadex LH-20)常用于忍冬苷的进一步纯化。聚酰胺对黄酮类化合物具有特异性吸附,分离效果较好。
- 制备型高效液相色谱(Prep-HPLC):对于高纯度忍冬苷的制备,Prep-HPLC是最有效的方法。通常采用C18反相色谱柱,以乙腈-水或甲醇-水为流动相,结合紫外检测器进行分离。
质量控制:
忍冬苷的鉴定和含量测定主要采用高效液相色谱(HPLC)和液相色谱-质谱联用(LC-MS)方法。紫外检测波长通常设定在254 nm或350 nm,以忍冬苷标准品进行定性和定量分析。此外,核磁共振波谱(NMR)和质谱(MS)用于结构确证。
忍冬苷在多种炎症模型中展现出显著的抗炎活性。在脂多糖(LPS)诱导的巨噬细胞炎症模型中,忍冬苷能够剂量依赖性地抑制促炎细胞因子如肿瘤坏死因子-α(TNF-α)、白细胞介素-6(IL-6)和白细胞介素-1β(IL-1β)的产生。同时,忍冬苷可降低一氧化氮(NO)和前列腺素E2(PGE2)的生成,这与抑制诱导型一氧化氮合酶(iNOS)和环氧合酶-2(COX-2)的表达有关。
在类风湿关节炎动物模型中,忍冬苷灌胃给药能够显著减轻胶原诱导关节炎(CIA)大鼠的关节肿胀、骨侵蚀和软骨破坏。组织病理学分析显示,忍冬苷治疗组关节滑膜组织中炎症细胞浸润减少,血管翳形成受到抑制。此外,忍冬苷可降低血清中类风湿因子(RF)和抗环瓜氨酸肽(CCP)抗体水平,提示其对自身免疫反应的调节作用。
忍冬苷的抗氧化活性源于其分子结构中的多个酚羟基。体外实验表明,忍冬苷能够有效清除1,1-二苯基-2-三硝基苯肼(DPPH)自由基、2,2′-联氮双(3-乙基苯并噻唑啉-6-磺酸)(ABTS)自由基和超氧阴离子自由基。其抗氧化能力与浓度呈正相关,在10-100 μM浓度范围内表现出显著的自由基清除活性。
在细胞氧化应激模型中,忍冬苷预处理可降低过氧化氢(H₂O₂)诱导的细胞活性氧(ROS)水平,提高超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-Px)和过氧化氢酶(CAT)的活性,同时增加还原型谷胱甘肽(GSH)含量。这些结果表明,忍冬苷通过直接清除自由基和增强内源性抗氧化防御系统发挥抗氧化作用。
忍冬苷对多种病原微生物具有抑制作用。研究发现,忍冬苷对铜绿假单胞菌(Pseudomonas aeruginosa)和白色念珠菌(Candida albicans)表现出显著的抗菌活性。其抗铜绿假单胞菌的机制部分归因于对藻酸盐分泌蛋白AlgE的抑制。AlgE是铜绿假单胞菌生物膜形成的关键蛋白,忍冬苷通过结合AlgE的活性位点,抑制藻酸盐的分泌,从而破坏生物膜的形成,增强细菌对宿主免疫防御和抗生素的敏感性。
对白色念珠菌的抑制作用涉及细胞膜完整性的破坏和菌丝形成的抑制。忍冬苷能够与真菌细胞膜中的麦角甾醇相互作用,增加膜通透性,导致细胞内物质外漏。同时,忍冬苷可下调菌丝特异性基因(如HWP1、ALS3)的表达,抑制酵母相向菌丝相的转变,从而降低真菌的致病性。
忍冬苷在神经退行性疾病模型中显示出保护作用。在谷氨酸诱导的神经元损伤模型中,忍冬苷预处理能够减轻神经元凋亡,降低乳酸脱氢酶(LDH)释放率。其神经保护机制与抑制氧化应激、减少细胞内钙超载和调节凋亡相关蛋白(Bax/Bcl-2比值)有关。
此外,忍冬苷可抑制β-淀粉样蛋白(Aβ)的聚集和纤维形成,这在阿尔茨海默病治疗中具有重要意义。通过结合Aβ单体,忍冬苷阻止其向毒性寡聚体和纤维的转化,同时促进已形成的Aβ纤维解聚。这些发现提示忍冬苷可能具有防治阿尔茨海默病的潜力。
忍冬苷在抗流感病毒方面展现出多靶点作用特点。研究表明,忍冬苷能够抑制甲型流感病毒(H1N1、H3N2亚型)在MDCK细胞中的复制,其半数抑制浓度(IC50)在微摩尔级别。作用机制涉及多个层面:
忍冬苷对黄嘌呤氧化酶(Xanthine Oxidase, XO)具有抑制作用,IC50为37.4 µg/mL(约62.9 μM)。XO是嘌呤代谢中的关键酶,催化次黄嘌呤和黄嘌呤转化为尿酸。XO过度激活导致尿酸生成增加,是痛风和高尿酸血症的重要病理机制。分子对接研究表明,忍冬苷的B环4′-羟基和C环羰基与XO的钼蝶呤辅因子活性位点形成氢键和π-π堆积相互作用,竞争性抑制底物与酶的结合。与临床常用的XO抑制剂别嘌醇相比,忍冬苷的抑制活性相对较弱,但其作为天然产物的安全性优势值得关注。
AlgE是铜绿假单胞菌外膜上的藻酸盐分泌通道蛋白,负责将藻酸盐从周质空间转运至细胞外。忍冬苷通过结合AlgE的孔道区域,阻断藻酸盐的分泌,从而抑制生物膜的形成。表面等离子体共振(SPR)实验证实,忍冬苷与AlgE的结合常数为微摩尔级别。这一发现为开发抗铜绿假单胞菌生物膜药物提供了新的先导化合物。
忍冬苷对流感病毒和宿主免疫系统的多靶点调控是其抗流感活性的核心机制:
NFE2L2(Nrf2)通路激活:忍冬苷可促进NFE2L2与Keap1的解离,使其核转位增加,进而结合抗氧化反应元件(ARE),启动下游抗氧化酶基因(如HO-1、NQO1)的转录。这一机制有助于减轻流感病毒感染引起的氧化应激损伤,保护宿主细胞。
NF-κB通路抑制:忍冬苷通过抑制IκBα的磷酸化和降解,阻止RELA(p65)和NFKB1(p50)的核转位,从而减少促炎细胞因子(TNF-α、IL-6、IL-1β)和趋化因子的表达。适度的NF-κB抑制有助于控制病毒感染引起的过度炎症反应,避免细胞因子风暴的发生。
MAPK8(JNK)信号调节:忍冬苷可抑制MAPK8(c-Jun N-terminal kinase)的磷酸化,调节细胞应激反应和凋亡信号。JNK通路的过度激活与流感病毒诱导的细胞凋亡和组织损伤密切相关,忍冬苷对该通路的抑制有助于保护肺组织完整性。
IRF3激活:忍冬苷可促进IRF3的磷酸化和二聚化,增强其核转位和转录活性,从而上调Ⅰ型干扰素(IFN-α/β)的表达。Ⅰ型干扰素是抗病毒天然免疫的关键效应分子,其诱导产生有助于抑制病毒复制和清除感染细胞。
直接抗病毒作用:忍冬苷与流感病毒表面蛋白HA和NA的结合,可阻断病毒的吸附和释放过程。此外,忍冬苷对病毒内部蛋白NP和PB2的相互作用,可干扰病毒基因组的复制和转录。
忍冬苷的作用机制呈现出复杂的信号网络调控特征。在流感病毒感染过程中,忍冬苷同时作用于病毒生命周期和宿主免疫反应,形成多层次的防御体系。通过激活NFE2L2抗氧化通路和IRF3干扰素通路,同时抑制NF-κB和MAPK8炎症通路,忍冬苷实现了对宿主免疫反应的精细调控——既增强抗病毒免疫,又避免过度炎症损伤。这种多靶点、多通路的调控模式,使得忍冬苷在抗流感治疗中具有独特的优势,可能降低病毒耐药性的产生风险。
基于计算药物化学和早期实验数据,忍冬苷的成药性特征如下:
综合来看,忍冬苷不符合Lipinski五规则中的多项标准,提示其口服生物利用度可能较低。然而,天然产物中许多成功的药物(如紫杉醇、环孢素)同样不符合Lipinski规则,因此不能仅凭此否定其成药性。近年来,针对黄酮苷类化合物的前药设计、纳米制剂和结构修饰策略,为改善其药代动力学性质提供了可能。
目前关于忍冬苷体内药代动力学的系统研究尚不充分,但基于其理化性质和同类化合物的研究,可推测以下特征:
为克服忍冬苷的药代动力学限制,可考虑以下策略:
忍冬苷作为多靶点抗流感先导化合物,具有以下独特优势:
然而,从先导化合物到临床药物仍面临诸多挑战:口服生物利用度的提升、抗病毒活性的优化、体内药效学和毒理学的系统评价等。未来可通过结构修饰和制剂技术,开发具有更好药代动力学性质的忍冬苷衍生物或类似物。
忍冬苷在类风湿关节炎治疗中的应用前景广阔。其抗炎机制涉及NF-κB和MAPK通路的调控,与当前临床使用的生物制剂(如TNF-α抑制剂)作用机制互补。此外,忍冬苷的抗氧化活性有助于减轻关节组织中的氧化应激损伤。开发忍冬苷作为辅助治疗药物,可能减少传统抗风湿药物的剂量和副作用。
忍冬苷对铜绿假单胞菌和白色念珠菌的抑制作用,特别是其抗生物膜活性,为治疗难治性感染提供了新思路。铜绿假单胞菌是医院获得性感染的主要病原体,其生物膜形成导致抗生素耐药和慢性感染。忍冬苷作为AlgE抑制剂,可与传统抗生素联合使用,增强抗感染效果。在抗真菌方面,忍冬苷对白色念珠菌菌丝形成的抑制作用,可用于治疗念珠菌感染,尤其是生物膜相关感染。
尽管忍冬苷血脑屏障穿透性较低,但其神经保护作用仍值得关注。通过前药设计或纳米递送系统,提高忍冬苷在中枢神经系统的分布,可能开发出治疗阿尔茨海默病、帕金森病等神经退行性疾病的候选药物。此外,忍冬苷的抗氧化和抗炎活性,使其在抗衰老和预防年龄相关疾病方面具有潜在应用价值。
忍冬苷作为一种天然黄酮苷类化合物,凭借其多样的生物活性和多靶点作用机制,在抗流感、抗炎、抗氧化、抗菌和神经保护等多个治疗领域展现出重要的研究价值。其独特的化学结构赋予其良好的水溶性和安全性特征,但也带来了口服生物利用度低等挑战。通过现代药物化学、药剂学和药理学手段,忍冬苷有望被开发成为治疗流感、类风湿关节炎和难治性感染的新型药物。
从传统中药金银花中的活性成分,到现代药物研发的先导化合物,忍冬苷的研究历程体现了天然产物药物发现的经典路径。未来,随着对忍冬苷作用机制认识的深入和制剂技术的进步,这一天然产物有望在临床应用中发挥更大的价值,为人类健康事业做出贡献。同时,忍冬苷的研究也为其他天然黄酮苷类化合物的开发提供了有益的借鉴和参考。

版权所有:© 成都普瑞法科技开发有限公司(2015)备案号:蜀ICP备15035167号-1 客服热线:400-829-7929
技术支持:南京库价















