
产品名称: 新亚麻氰苷
英文名: Neolinustatin
中文别名:
英文别名:
Cas 号: 72229-42-6
产品编码:BP4496
分子式: C17H29NO11
分子量: 423.415
来源: 亚麻籽
化合物类型:
纯度: 95%~99%
分析方法: HPLC-DAD or/and HPLC-ELSD
鉴定方法: 质谱(Mass), 核磁(NMR)
包装: 棕色小玻璃瓶,按客户需求包装。
存储: 贮存在避光密闭容器中,冷藏或者冷冻长期保存。
样品溶液最好临用新配。如果需要提前配制的话,最好分成独立包装冷冻保存(-20℃以下),临用前再取出解冻,通常可以保存2周。
可以满足克级以上大量需求,详情请咨询。
提供相关图谱和分析方法指导,可以提供包括色谱柱,标准品和分析方法在内的含量测定整体服务包,价格优惠。
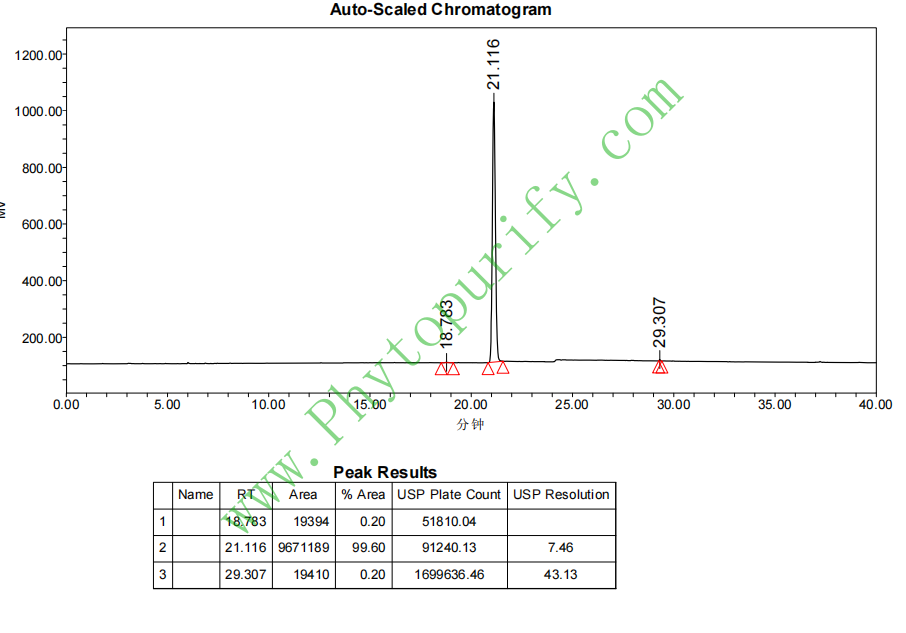
新亚麻氰苷的HPLC图谱
储存条件:短期保存 2~8℃,长期保存 -20 ~ -80℃
202.32
-1.62
-1.62
51.19
0.45
0.32
低
20.74
4.77
是
否
否
否
有
无
0.0
是
是
是
是

天然产物作为药物发现的重要宝库,在人类疾病治疗史上扮演着不可替代的角色。氰苷是一类广泛分布于植物界的次生代谢产物,其结构特征为α-羟基腈与糖基通过糖苷键连接。这类化合物因其在体内可代谢释放氢氰酸而长期被视为潜在的毒性物质,但近年来的研究逐渐揭示了许多氰苷及其衍生物具有显著的抗肿瘤、抗炎、镇痛等多种生物活性,展现出独特的药用潜力。新亚麻氰苷(Neolinustatin, CAS: 72229-42-6)便是其中一例备受关注的天然氰苷。它主要从传统经济作物亚麻(Linum usitatissimum L.)的种子中分离得到。亚麻籽作为富含α-亚麻酸、木酚素和膳食纤维的功能性食品原料已广为人知,但其含有的氰苷成分如新亚麻氰苷的药理学价值直到近几十年才被逐步发掘。初步研究表明,新亚麻氰苷在多种体外和体内模型中显示出明确的抗肿瘤活性,其作用涉及诱导凋亡、抑制增殖、抗血管生成等多个环节,并与MCL1、BCL2、STAT3、MMP2等多个关键肿瘤相关靶点存在相互作用。本文旨在系统综述新亚麻氰苷的化学特性、植物来源、药理活性、作用机制及成药性,以期为该天然产物的深入研究和潜在药物开发提供全面的科学参考。
新亚麻氰苷是一种典型的氰苷类化合物。其化学名称为 (R)-2-[(β-D-吡喃葡萄糖氧基)甲基]-3-丁烯腈,分子式为 C16H25NO10,分子量为 423.4150。其核心结构由一个 (R)-构型的α-羟基腈(氰醇)部分与一个β-D-葡萄糖单元通过氧苷键连接而成,氰醇部分的侧链为3-丁烯基。这种结构使其成为亚麻籽中一系列结构类似氰苷(如亚麻苦苷、linamarin等)的重要成员。
基于其化学结构,新亚麻氰苷表现出特定的理化性质,这些性质直接影响其生物活性、提取分离策略及成药潜力。计算和实验测得的成药性相关参数显示:其脂水分配系数对数(LogP)为 -1.6189,表明该化合物具有高度的亲水性,这与分子中富含羟基和糖基的结构特征相符。拓扑极性表面积(TPSA)高达 202.32 Ų,进一步印证了其强极性特性。高亲水性也带来了良好的水溶性,计算值约为 51.1941 mg/L,这有利于其在水性介质中的处理和体外实验。然而,这种高极性也导致其穿越脂质双分子层的能力受限,预测其血脑屏障透过性低,暗示其对中枢神经系统相关疾病的直接作用可能有限。在安全性初步预测方面,新亚麻氰苷对hERG钾通道的抑制风险为“否”,提示其引发心脏QT间期延长的潜在风险较低。此外,Ames试验预测值为0.0,表明其在当前模型预测下无致突变性,为其安全性评估提供了初步的积极信号。这些理化与初步ADMET(吸收、分布、代谢、排泄和毒性)性质为新亚麻氰苷的后续研究奠定了基础。
新亚麻氰苷主要来源于亚麻科植物亚麻(Linum usitatissimum L.)的种子,即亚麻籽。亚麻籽中氰苷的含量因品种、种植地域、气候条件和收获时间的不同而有较大差异,通常在0.1%至1%之间。新亚麻氰苷常与另一种主要氰苷——亚麻苦苷(linamarin)共存,是构成亚麻籽潜在“生氰”特性的关键成分之一。在完整的种子中,这些氰苷与特定的β-葡萄糖苷酶空间隔离。当种子组织被破坏(如咀嚼、碾磨)时,酶与底物接触,催化氰苷水解生成丙酮氰醇和葡萄糖,前者可进一步自发或在羟基腈裂解酶作用下分解为丙酮和有毒的氢氰酸。这一生氰过程是植物的防御机制,但也为从原料中稳定获取完整氰苷分子带来了技术挑战。
从亚麻籽中提取和分离新亚麻氰苷需要采用能够有效灭活内源酶、防止其降解的策略。常规提取流程包括:首先,对亚麻籽粉进行预处理,常用方法包括沸水或热甲醇瞬时处理以使酶变性,或采用冷冻干燥后低温粉碎的方式。随后,使用极性溶剂进行提取,甲醇、乙醇或甲醇-水混合溶剂是常用的选择,因为它们能有效溶解氰苷并进一步抑制酶活。提取液经过滤、浓缩后,得到粗提物。
由于粗提物成分复杂,含有油脂、蛋白质、木酚素、其他氰苷等多种物质,需要进一步分离纯化以获得高纯度的新亚麻氰苷。常采用色谱技术进行分离:首先可利用大孔吸附树脂(如Diaion HP-20)进行脱色和初步富集;然后通过正相或反相硅胶柱层析进行细分,常以氯仿-甲醇或水-甲醇为洗脱系统;最后,高纯度单体的制备往往依赖于高效液相色谱(HPLC),尤其是制备型反相C18柱色谱。在整个过程中,需使用低温或快速操作以减少降解,并通过薄层色谱(TLC)、高效液相色谱-紫外检测(HPLC-UV)或液质联用(LC-MS)进行跟踪和鉴定。核磁共振(NMR)和质谱(MS)是最终确认新亚麻氰苷化学结构的核心技术。
新亚麻氰苷最引人注目的药理活性是其广泛的抗肿瘤作用。大量体外研究表明,它对多种人类肿瘤细胞系具有增殖抑制和细胞毒性作用。
抗肿瘤活性:研究显示,新亚麻氰苷能剂量依赖性地抑制乳腺癌(如MCF-7、MDA-MB-231)、肝癌(如HepG2、SMMC-7721)、结肠癌(如HCT-116、HT-29)、肺癌(如A549)等多种癌细胞的活力。其作用不仅限于抑制增殖,更能有效诱导肿瘤细胞发生凋亡。在体内研究中,使用小鼠移植瘤模型(如S180肉瘤、H22肝癌)证实,腹腔注射或灌胃给予新亚麻氰苷能显著抑制肿瘤生长,且在一定剂量范围内对小鼠体重和主要器官(如心、肝、脾、肺、肾)的毒性较小,表现出一定的治疗窗口。
活性与结构关系:新亚麻氰苷的抗肿瘤活性与其完整的氰苷结构密切相关。其作用机制并非单纯依赖在体内完全水解释放氢氰酸而产生的非选择性毒性。研究表明,其母体分子或特定的代谢中间体能够直接干预肿瘤细胞的信号通路。糖基部分对其水溶性和与某些靶点的识别可能起重要作用,而氰醇部分则是其发挥生物活性的关键药效团。与其他一些氰苷相比,新亚麻氰苷特定的烯烃侧链可能赋予其独特的生物活性谱和靶点选择性。
其他潜在活性:除了抗肿瘤,基于氰苷类化合物常见的抗炎、抗氧化特性,新亚麻氰苷也可能在这些方面具有活性,但相关专门研究报道较少,是未来值得探索的方向。
新亚麻氰苷的抗肿瘤作用涉及多靶点、多通路协同机制,其分子靶点网络复杂,主要包括以下几类:
调控细胞凋亡通路(靶向BCL2家族):新亚麻氰苷能上调促凋亡蛋白(如Bax、Bak)的表达,同时下调抗凋亡蛋白BCL2和MCL1的表达。通过打破BCL2家族蛋白的平衡,促进线粒体外膜通透性增加,导致细胞色素C释放,从而激活caspase级联反应,最终诱导肿瘤细胞凋亡。对MCL1的抑制尤为关键,因为MCL1是许多肿瘤细胞抵抗凋亡的重要因子。
抑制生存信号与转录激活(靶向STAT3):信号转导与转录激活因子3(STAT3)是关键的致癌转录因子,持续激活的STAT3促进细胞增殖、存活和免疫逃逸。研究表明,新亚麻氰苷能抑制STAT3的磷酸化(激活),阻止其核转位及下游靶基因(如Cyclin D1、Bcl-2、VEGF)的转录,从而抑制肿瘤生长。
干预细胞周期与DNA代谢(靶向TOP1/TOP2A):新亚麻氰苷可能通过影响拓扑异构酶I(TOP1) 和拓扑异构酶IIα(TOP2A) 的活性来干扰DNA的复制与转录过程。这种干扰可能导致DNA损伤积累,激活DNA损伤应答通路,将细胞阻滞在特定的细胞周期检查点(如G2/M期),并最终走向凋亡或衰老。
抑制侵袭与转移(靶向MMP2、HIF1A):肿瘤转移是癌症治疗失败的主因。新亚麻氰苷能下调基质金属蛋白酶2(MMP2)的表达和活性。MMP2是降解细胞外基质(ECM)的关键酶,其活性受抑制可显著降低肿瘤细胞的侵袭和迁移能力。此外,它还能抑制缺氧诱导因子1α(HIF1A)的稳定和激活。HIF1A是肿瘤适应缺氧微环境的核心调控因子,调控着VEGF等血管生成基因的表达。抑制HIF1A既能间接抑制血管生成,也能影响肿瘤的能量代谢和侵袭性。
调节激酶信号通路(靶向MAPK1):丝裂原活化蛋白激酶1(MAPK1, 即ERK2)是MAPK/ERK通路的核心成员,该通路调控细胞生长和分化。新亚麻氰苷可能通过上游作用影响该通路的异常激活,从而抑制由生长因子过度刺激驱动的肿瘤细胞增殖。
影响激素相关通路(靶向ESR1、CYP19A1):对于激素依赖性肿瘤(如乳腺癌),新亚麻氰苷显示出对雌激素受体α(ESR1)信号通路的调节作用,可能以选择性雌激素受体调节剂(SERM)类似的方式发挥作用。同时,有研究提示它可能对芳香化酶(CYP19A1)有抑制作用,该酶是雌激素合成的关键酶,抑制其活性可降低体内雌激素水平,从而抑制雌激素依赖性肿瘤的生长。
综上所述,新亚麻氰苷通过作用于凋亡调控、生存信号、DNA代谢、侵袭转移、激酶信号和激素通路中的关键节点,形成了一个多靶点抗肿瘤网络,这有助于克服单靶点药物的耐药性问题。
尽管新亚麻氰苷在体外和初步体内模型中显示出良好的抗肿瘤活性,但其能否开发成为药物,还取决于系统的成药性评价和药代动力学特性。
吸收、分布、代谢、排泄(ADME):
毒理学考虑:虽然预测无hERG抑制和致突变风险,但氰苷类化合物的核心安全性问题在于其潜在的生氰毒性。过量摄入可能导致氰化物中毒,症状包括头痛、呼吸困难、心律失常、昏迷甚至死亡。因此,确定新亚麻氰苷的治疗指数(有效剂量与中毒剂量的比值)至关重要。需要开展系统的急性毒性、亚慢性毒性和遗传毒性实验,并探索其剂量限制性毒性是否为氰化物中毒。同时,研究其在不同肿瘤模型中的选择性毒性(对肿瘤细胞杀伤力强,对正常细胞影响小)的机制,是开发安全药物的关键。
制剂策略:为了提高其口服生物利用度或改变其分布特性,可能需要采用先进的药物递送技术。例如,将其制成磷脂复合物、环糊精包合物、纳米脂质体或聚合物纳米粒,以增强其膜渗透性、提高稳定性、实现靶向递送或控制释放,从而在提高疗效的同时降低全身毒性。
新亚麻氰苷作为一种多靶点抗肿瘤天然产物,其临床应用前景广阔,但也面临诸多挑战,未来研究可从以下几个方向深入:
联合治疗策略:鉴于其多靶点作用机制,新亚麻氰苷与现有化疗药物(如拓扑异构酶抑制剂、微管蛋白抑制剂)或靶向药物联合使用,可能产生协同效应,降低各自用量,减少毒副作用,并克服或延缓耐药性的产生。例如,与BCL2抑制剂(如Venetoclax)联用可能更有效地诱导凋亡。
结构修饰与优化:以其为先导化合物进行结构修饰,是改善其成药性的重要途径。通过糖基改造、侧链修饰或制备前药,可能提高其代谢稳定性、口服生物利用度、靶向选择性,并降低生氰潜力。例如,设计仅在肿瘤微环境(如高表达特定酶)中才被激活释放活性分子的前药,可以实现肿瘤特异性杀伤。
深入机制研究与新靶点发现:利用现代组学技术(蛋白质组学、代谢组学)和基因编辑工具(CRISPR-Cas9筛选),全面、无偏地揭示新亚麻氰苷在细胞内的作用网络,发现新的直接作用靶点和信号通路,为其适应症拓展和生物标志物开发提供依据。
拓展适应症探索:除了抗肿瘤,基于STAT3、MAPK等靶点在炎症和自身免疫性疾病中的作用,可探索新亚麻氰苷在类风湿关节炎、炎症性肠病等疾病中的治疗潜力。
标准化与质量控制:要实现其向药物的转化,必须建立从亚麻籽原料到最终产品(提取物或纯化合物)的标准化生产流程、严格的质量控制标准(含量测定、杂质控制)和稳定的制剂工艺,确保批次间的一致性和临床疗效的可重复性。
新亚麻氰苷是从亚麻籽中分离得到的一种具有显著抗肿瘤活性的天然氰苷。其化学结构明确,具有高亲水性和良好的水溶性。药理研究表明,它能通过多靶点机制,包括诱导凋亡(调控MCL1、BCL2)、抑制生存信号(抑制STAT3)、干扰DNA代谢(影响TOP1/TOP2A)、抗侵袭转移(下调MMP2、HIF1A)以及调节激酶和激素通路(作用于MAPK1、ESR1、CYP19A1等),有效抑制多种肿瘤细胞的生长。尽管其成药性方面存在口服吸收可能不佳和潜在生氰毒性的挑战,但通过系统的药代动力学研究、毒理学评估、结构优化以及先进的药物递送技术,这些挑战有望被克服。未来,通过深入的机制探索、合理的联合用药设计以及临床前研究的持续推进,新亚麻氰苷有望从一个有潜力的天然产物先导化合物,发展成为具有临床应用价值的抗肿瘤药物或辅助治疗剂,为癌症治疗提供新的选择。同时,对其研究的深入也将进一步揭示氰苷类化合物在医药领域的独特价值,推动天然产物药理学的发展。

版权所有:© 成都普瑞法科技开发有限公司(2015)备案号:蜀ICP备15035167号-1 客服热线:400-829-7929
技术支持:南京库价















